Ch 10 STUDY GUIDE
Sensory Systems
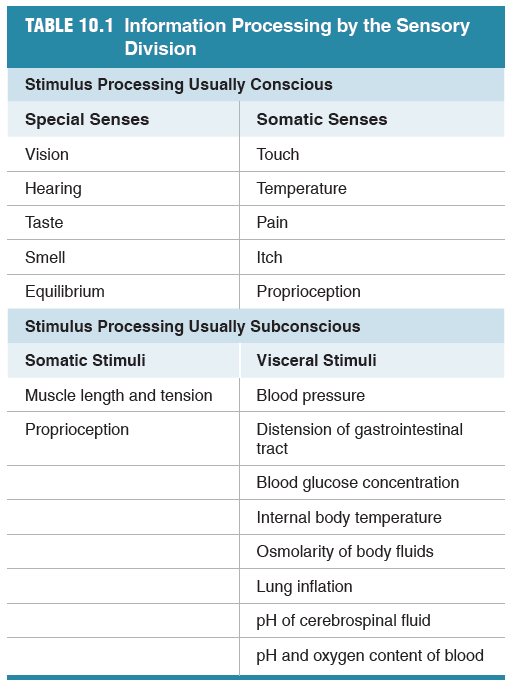
Some senses are called "special senses". These are vision, hearing, taste and smell. We also have "somatic senses" which include touch, temperature, pain, itch and proprioception.
Proprioception, which is defined as the awareness of body
movement and position in space, is mediated by muscle and joint
sensory receptors called proprioceptors and may be either
unconscious or conscious. If you close your eyes and raise your
arm above your head, you are aware of the arm’s position because
of proprioceptor activation.
Proprioception, which is defined as the awareness of body
movement and position in space, is mediated by muscle and joint
sensory receptors called proprioceptors and may be either
unconscious or conscious. If you close your eyes and raise your
arm above your head, you are aware of the arm’s position because
of proprioceptor activation.
10.1 General Properties of Sensory Systems
All sensory pathways have certain elements in common. They
begin with a stimulus, in the form of physical energy that acts
on a sensory receptor. The receptor, or sensor, is a transducer that
converts the stimulus into an intracellular signal, which is usually
a change in membrane potential. If the stimulus is above threshold,
action potentials pass along a sensory neuron to the central nervous
system (CNS), where incoming signals are integrated. Some stimuli
pass upward to the cerebral cortex, where they reach conscious
perception, but others are acted on subconsciously, without our
awareness. At each synapse along the pathway, the nervous system
can modulate and shape the sensory information.
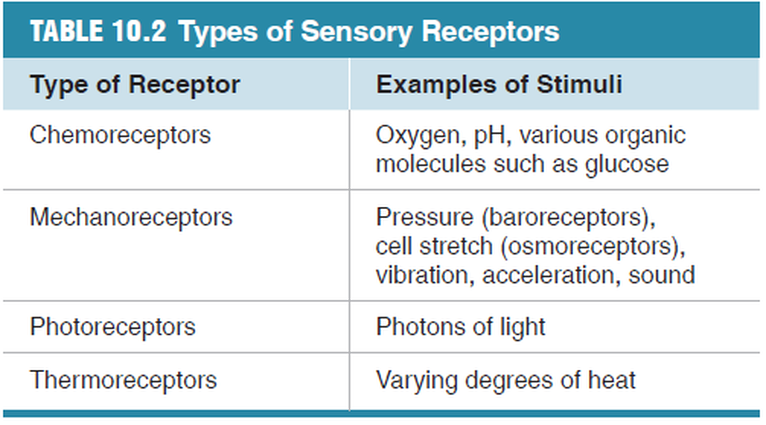
Receptors Are Sensitive to Particular Forms of Energy
Receptors can be divided into four major groups, based on
the type of stimulus to which they are most sensitive.
Chemoreceptors respond to chemical ligands that bind to the
receptor (taste and smell, for example). Mechanoreceptors
respond to various forms of mechanical energy, including pressure,
vibration, gravity, acceleration, and sound (hearing, for
example). Thermoreceptors respond to temperature, and
photoreceptors for vision respond to light.
the type of stimulus to which they are most sensitive.
Chemoreceptors respond to chemical ligands that bind to the
receptor (taste and smell, for example). Mechanoreceptors
respond to various forms of mechanical energy, including pressure,
vibration, gravity, acceleration, and sound (hearing, for
example). Thermoreceptors respond to temperature, and
photoreceptors for vision respond to light.
A Sensory Neuron Has a Receptive Field
Somatic sensory and visual neurons are activated by stimuli that fall
within a specific physical area known as the neuron’s receptive
field.
Receptive fields frequently overlap with neighboring
receptive fields. In addition, sensory neurons of neighboring receptive fields
may exhibit convergence, in which multiple presynaptic neurons
provide input to a smaller number of postsynaptic neurons.
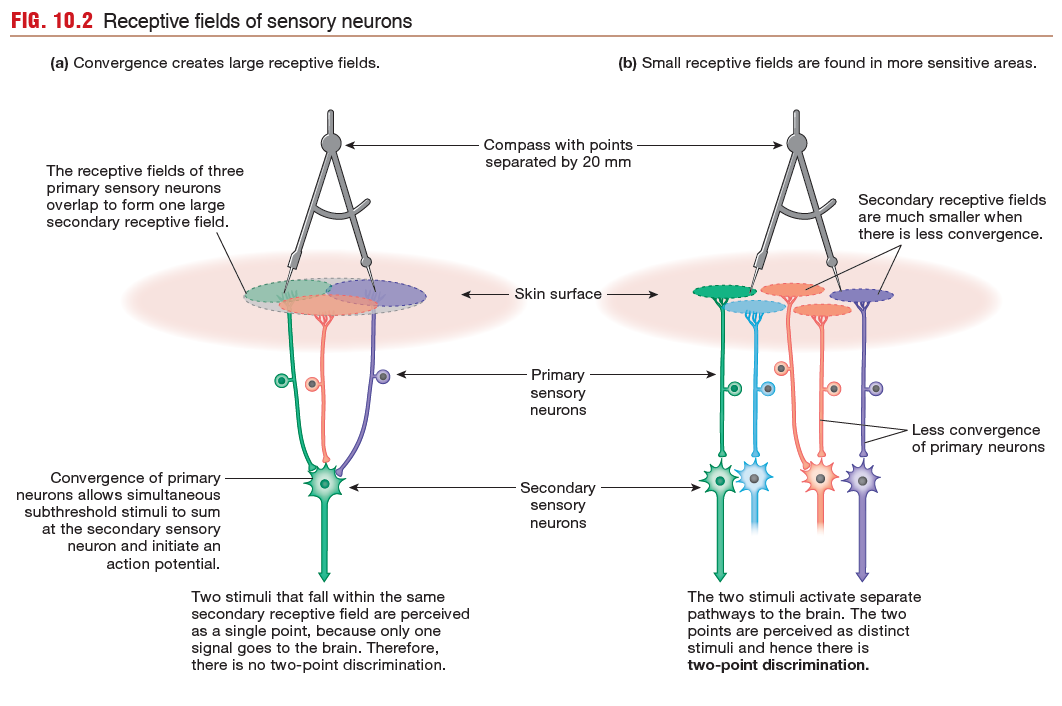
The size of secondary receptive fields determines how sensitive
a given area is to a stimulus. For example, sensitivity to touch
is demonstrated by a two-point discrimination test. In some
regions of skin, such as that on the arms and legs, two pins placed
within 20 mm of each other are interpreted by the brain as a single
pinprick. In these areas, many primary neurons converge on a
single secondary neuron, so the secondary receptive field is very
large (Fig. 10.2a).
In contrast, more sensitive areas of skin, such as the fingertips,
have smaller receptive fields, with as little as a 1:1 relationship
between primary and secondary sensory neurons (Fig. 10.2b). In
these regions, two pins separated by as little as 2 mm can be perceived
as two separate touches.
a given area is to a stimulus. For example, sensitivity to touch
is demonstrated by a two-point discrimination test. In some
regions of skin, such as that on the arms and legs, two pins placed
within 20 mm of each other are interpreted by the brain as a single
pinprick. In these areas, many primary neurons converge on a
single secondary neuron, so the secondary receptive field is very
large (Fig. 10.2a).
In contrast, more sensitive areas of skin, such as the fingertips,
have smaller receptive fields, with as little as a 1:1 relationship
between primary and secondary sensory neurons (Fig. 10.2b). In
these regions, two pins separated by as little as 2 mm can be perceived
as two separate touches.
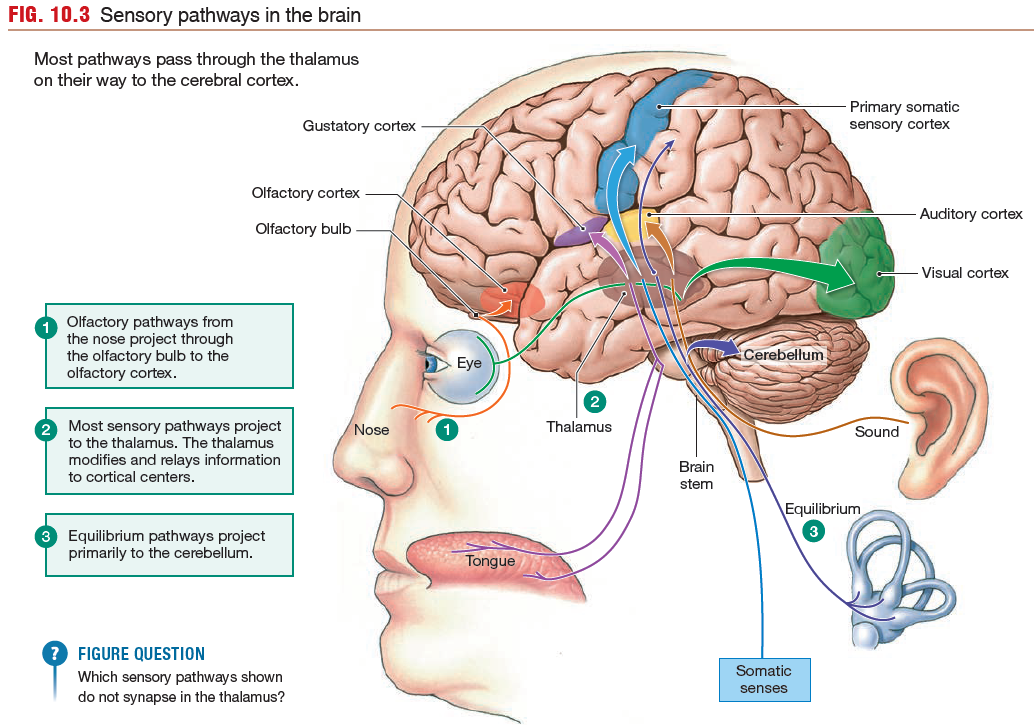
The CNS Integrates Sensory Information
Sensory information from much of the body enters the spinal
cord and travels through ascending pathways to the brain. Each major division of the brain processes one or more types of sensory information.
Coding and Processing Distinguish
Stimulus Properties
The CNS must distinguish four properties of a stimulus:
(1) its nature, or modality
(2) its location
(3) its intensity
(4) its duration
Sensory Modality
The modality of a stimulus is indicated by which
sensory neurons are activated and by where the pathways of the
activated neurons terminate in the brain.
sensory neurons are activated and by where the pathways of the
activated neurons terminate in the brain.
Location of the Stimulus
The location of a stimulus is also
coded according to which receptive fields are activated.
For example, touch receptors in the hand project to a specific area of the cerebral cortex. Similarly, one type of the phantom limb pain reported by amputees occurs when secondary sensory neurons in the spinal cord become hyperactive, resulting in the sensation of pain in a limb that is no longer there.
coded according to which receptive fields are activated.
For example, touch receptors in the hand project to a specific area of the cerebral cortex. Similarly, one type of the phantom limb pain reported by amputees occurs when secondary sensory neurons in the spinal cord become hyperactive, resulting in the sensation of pain in a limb that is no longer there.
Auditory information is an exception to the localization rule,
however. The brain uses the timing of receptor activation to compute a location,
as shown in FIGURE.
however. The brain uses the timing of receptor activation to compute a location,
as shown in FIGURE.
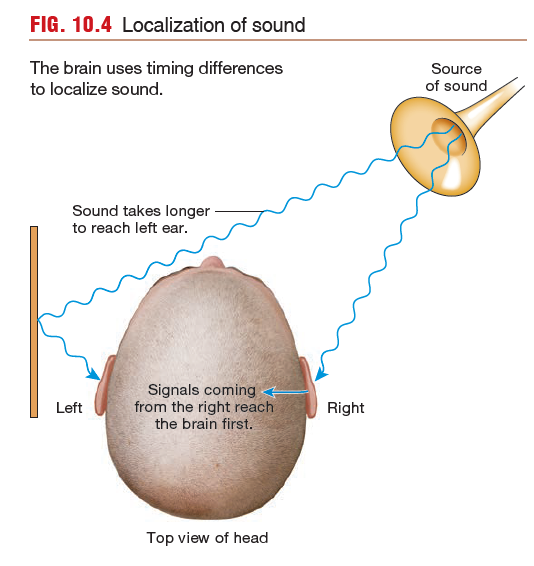
A sound originating directly in front of a person reaches both
ears simultaneously. A sound originating on one side reaches the
closer ear several milliseconds before it reaches the other ear. The
brain registers the difference in the time it takes for the sound
stimuli to reach the two sides of the auditory cortex and uses that
information to compute the sound’s source.
Intensity of the Stimulus
The intensity of a stimulus is coded in two types of information:
Population coding for intensity occurs because the threshold
for the preferred stimulus is not the same for all receptors. Only the
most sensitive receptors (those with the lowest thresholds) respond
to a low-intensity stimulus. As a stimulus increases in intensity,
additional receptors are activated. The CNS then translates the
number of active receptors into a measure of stimulus intensity.
- the number of receptors activated (population coding)
- the frequency of action potentials (frequency coding)
Population coding for intensity occurs because the threshold
for the preferred stimulus is not the same for all receptors. Only the
most sensitive receptors (those with the lowest thresholds) respond
to a low-intensity stimulus. As a stimulus increases in intensity,
additional receptors are activated. The CNS then translates the
number of active receptors into a measure of stimulus intensity.
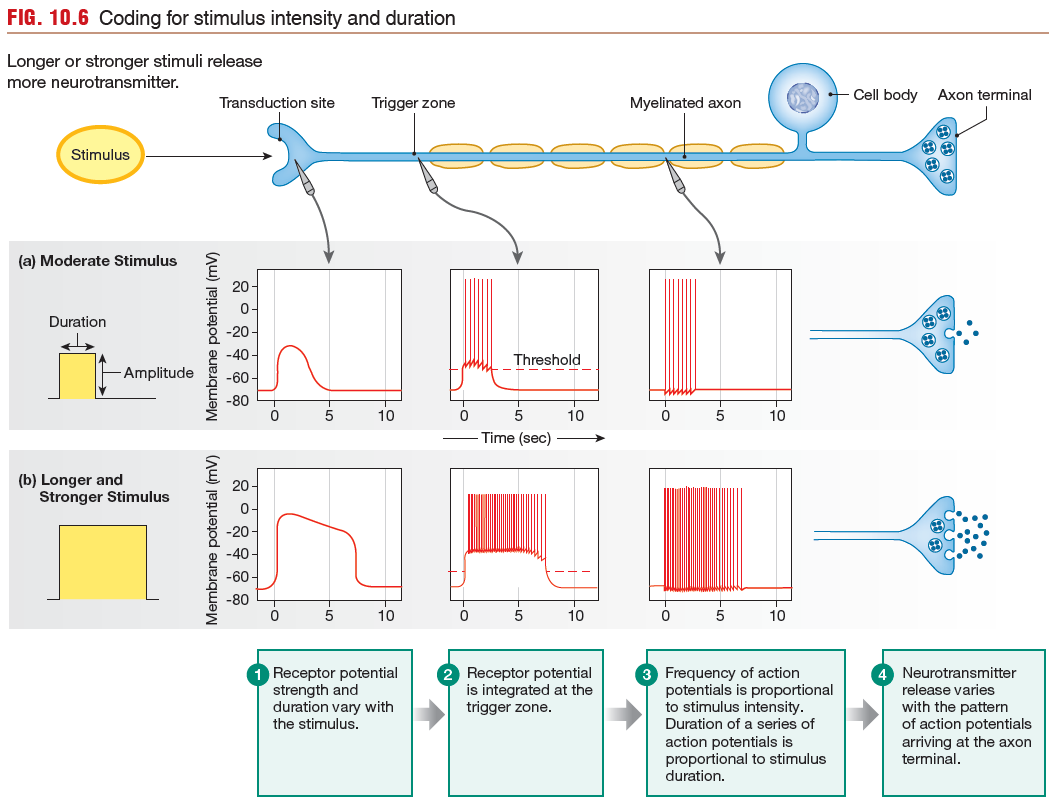
Duration of the Stimulus
The duration of a stimulus is coded by
the duration of action potentials in the sensory neuron. In general,
a longer stimulus generates a longer series of action potentials in
the primary sensory neuron.
the duration of action potentials in the sensory neuron. In general,
a longer stimulus generates a longer series of action potentials in
the primary sensory neuron.
The specificity of sensory pathways is established in several ways:
- Each receptor is most sensitive to a particular type of stimulus.
- A stimulus above threshold initiates action potentials in a sensory neuron that projects to the CNS.
- Stimulus intensity and duration are coded in the pattern of action potentials reaching the CNS.
- Stimulus location and modality are coded according to which receptors are activated or (in the case of sound) by the timing of receptor activation.
- Each sensory pathway projects to a specific region of the cerebral cortex dedicated to a particular receptive field. The brain can then tell the origin of each incoming signal.
10.2 Somatic Senses
There are four somatosensory modalities:
- touch
- proprioception
- temperature
- nociception (pain and itch)
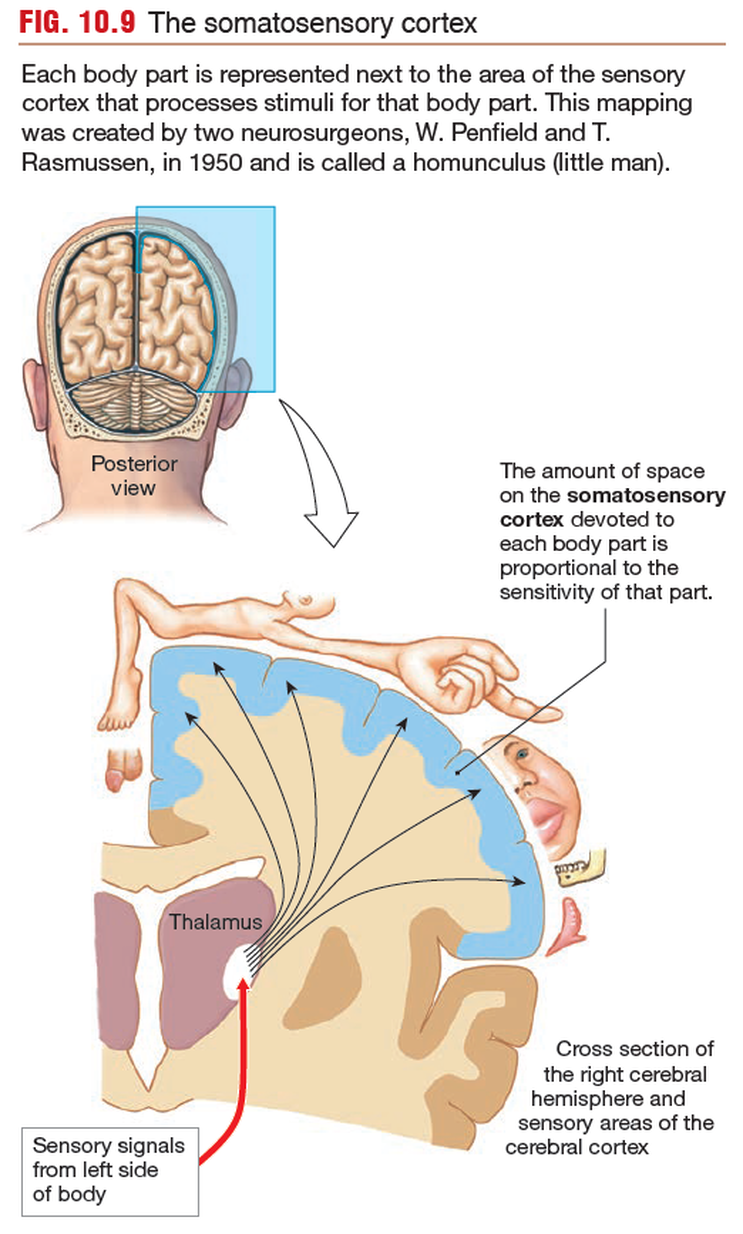
The somatosensory cortex
The somatosensory cortex is the part of the brain
that recognizes where ascending sensory tracts originate. Each sensory
tract has a corresponding region of the cortex, its sensory field.
All sensory pathways for the left hand terminate in one area, all
pathways for the left foot terminate in another area, and so on.
Within the cortical region for a particular body part,
columns of neurons are devoted to particular types of receptors.
For example, a cortical column activated by cold receptors
in the left hand may be found next to a column activated by
pressure receptors in the skin of the left hand. This columnar
arrangement creates a highly organized structure that maintains
the association between specific receptors and the sensory modality
they transmit.
that recognizes where ascending sensory tracts originate. Each sensory
tract has a corresponding region of the cortex, its sensory field.
All sensory pathways for the left hand terminate in one area, all
pathways for the left foot terminate in another area, and so on.
Within the cortical region for a particular body part,
columns of neurons are devoted to particular types of receptors.
For example, a cortical column activated by cold receptors
in the left hand may be found next to a column activated by
pressure receptors in the skin of the left hand. This columnar
arrangement creates a highly organized structure that maintains
the association between specific receptors and the sensory modality
they transmit.
Touch Receptors Respond to Many Different Stimuli
Touch receptors are among the most common receptors in the
body. These receptors respond to many forms of physical contact, such as stretch, steady pressure, fluttering or stroking movement, vibration, and texture. They are found both in the skin and in deeper regions of the body.
Pacinian corpuscles respond best to high-frequency vibrations,
whose energy is transferred through the connective tissue capsule
to the nerve ending, where the energy opens mechanically gated
ion channels.
The Merkel receptor consists of a Merkel cell
that synapses onto a primary sensory neuron. Merkel receptors are
found in the greatest density in the fingertips and are responsible
for the high sensitivity of tactile reception in these areas.
Skin Temperature Receptors Are Free Nerve Endings.
Temperature receptors, or thermoreceptors, are found throughout the
body in skin, muscles, internal organs, and the CNS because of the
importance of temperature homeostasis. In the skin, thermoreceptors
are free nerve endings that terminate in the subcutaneous layers.
Cold receptors are sensitive primarily to temperatures lower
than body temperature. Warm receptors are stimulated by temperatures
in the range extending from normal body temperature
(37 °C) to about 45 °C. Above that temperature, pain receptors are
activated, creating a sensation of painful heat.
body. These receptors respond to many forms of physical contact, such as stretch, steady pressure, fluttering or stroking movement, vibration, and texture. They are found both in the skin and in deeper regions of the body.
Pacinian corpuscles respond best to high-frequency vibrations,
whose energy is transferred through the connective tissue capsule
to the nerve ending, where the energy opens mechanically gated
ion channels.
The Merkel receptor consists of a Merkel cell
that synapses onto a primary sensory neuron. Merkel receptors are
found in the greatest density in the fingertips and are responsible
for the high sensitivity of tactile reception in these areas.
Skin Temperature Receptors Are Free Nerve Endings.
Temperature receptors, or thermoreceptors, are found throughout the
body in skin, muscles, internal organs, and the CNS because of the
importance of temperature homeostasis. In the skin, thermoreceptors
are free nerve endings that terminate in the subcutaneous layers.
Cold receptors are sensitive primarily to temperatures lower
than body temperature. Warm receptors are stimulated by temperatures
in the range extending from normal body temperature
(37 °C) to about 45 °C. Above that temperature, pain receptors are
activated, creating a sensation of painful heat.
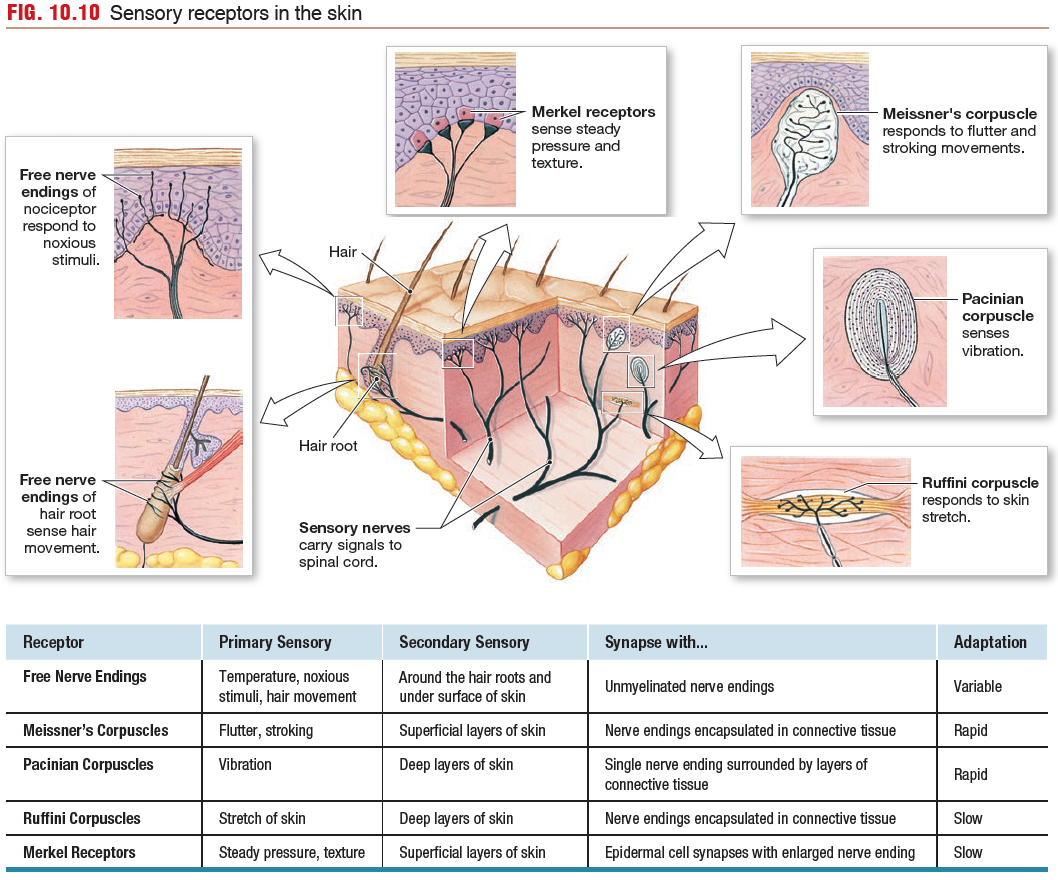
Nociceptors Initiate Protective Responses. Nociceptors are neurons with free nerve endings that respond to a variety of strong harmful
stimuli (chemical, mechanical, or thermal) that cause or have the
potential to cause tissue damage.
Nociceptors are found in the skin, joints, muscles, bones and various internal organs, but not in the central nervous system.
Afferent signals from nociceptors are carried to the CNS in two types of primary sensory fibers:
ITCH - When histamine or some other stimulus activates a subtype of C fiber, we
perceive the sensation we call itch.
PAIN - Pain is a subjective perception, the brain’s interpretation of sensory information transmitted along pathways that begin at nociceptors.
The timing distinction between the two is most obvious when the stimulus originates far from the CNS, such as when you stub your toe. You first experience a quick stabbing sensation (fast pain), followed shortly by a dull throbbing (slow pain). Itch (pruritus) comes only from nociceptors in the skin and is characteristic of many rashes and other skin conditions.
stimuli (chemical, mechanical, or thermal) that cause or have the
potential to cause tissue damage.
Nociceptors are found in the skin, joints, muscles, bones and various internal organs, but not in the central nervous system.
Afferent signals from nociceptors are carried to the CNS in two types of primary sensory fibers:
- A delta fibers
- C fibers
ITCH - When histamine or some other stimulus activates a subtype of C fiber, we
perceive the sensation we call itch.
PAIN - Pain is a subjective perception, the brain’s interpretation of sensory information transmitted along pathways that begin at nociceptors.
- Fast pain, described as sharp and localized, is rapidly transmitted to the CNS by the small, myelinated Ad fibers.
- Slow pain, described as duller and more diffuse, is carried on small, unmyelinated C fibers.
The timing distinction between the two is most obvious when the stimulus originates far from the CNS, such as when you stub your toe. You first experience a quick stabbing sensation (fast pain), followed shortly by a dull throbbing (slow pain). Itch (pruritus) comes only from nociceptors in the skin and is characteristic of many rashes and other skin conditions.

10.3 Chemoreception: Smell and Taste
Smell and taste are both forms of chemoreception,
one of the oldest senses from an evolutionary perspective. Olfaction allows us to discriminate among millions of different odors. Even so, our noses are not nearly as sensitive as those of many other animals whose survival depends on olfactory
cues. The olfactory bulb, the extension of the forebrain that
receives input from the primary olfactory neurons, is much better
developed in vertebrates whose survival is more closely linked to
chemical monitoring of their environment (
one of the oldest senses from an evolutionary perspective. Olfaction allows us to discriminate among millions of different odors. Even so, our noses are not nearly as sensitive as those of many other animals whose survival depends on olfactory
cues. The olfactory bulb, the extension of the forebrain that
receives input from the primary olfactory neurons, is much better
developed in vertebrates whose survival is more closely linked to
chemical monitoring of their environment (
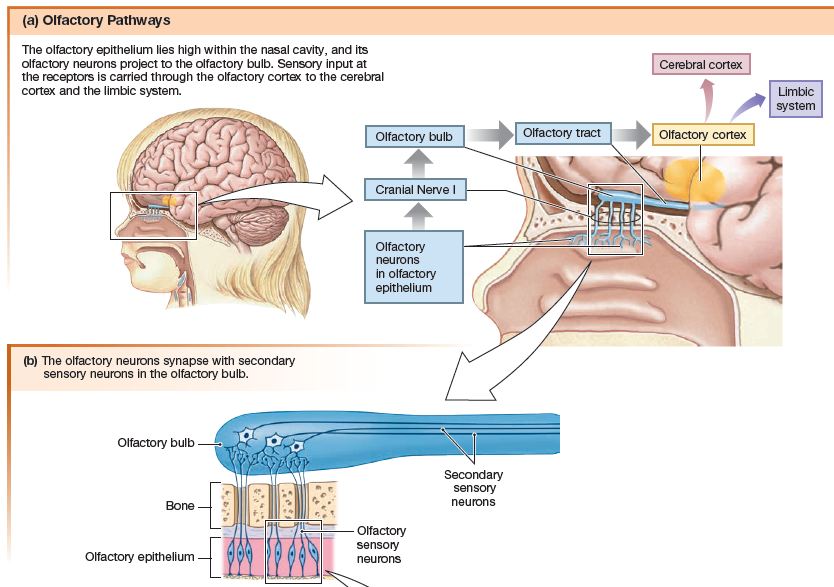
Olfactory Pathways
The human olfactory system consists of an
olfactory epithelium lining the nasal cavity, with embedded primary
sensory neurons called olfactory sensory neurons. Axons of
the olfactory sensory neurons form the olfactory nerve, or cranial
nerve I. The olfactory nerve synapses with secondary sensory
neurons in the olfactory bulb, located on the underside of the
frontal lobe. Secondary and higher-order neurons
project from the olfactory bulb through the olfactory tract to the
olfactory cortex. The olfactory tract, unlike most other
sensory pathways, bypasses the thalamus.
Ascending pathways from the olfactory bulb also lead to the
amygdala and hippocampus, parts of the limbic system involved
with emotion and memory. The link between smell, memory, and
emotion is one amazing aspect of olfaction.
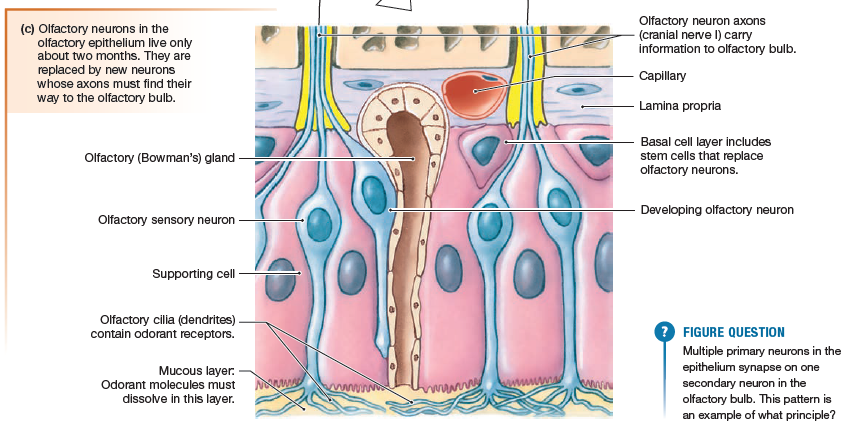
Olfactory Signal Transduction
The surface of the olfactory epithelium
is composed of the olfactory sensory neuron
dendrites, each knob branching into multiple nonmotile cilia. The cilia are embedded in a layer of mucus that is produced
by olfactory (Bowman’s) glands in the epithelium and basal lamina.
Odorant molecules must first dissolve in and penetrate the mucus before they can
bind to an olfactory receptor protein on the olfactory cilia. Each
olfactory receptor is sensitive to a limited range of odorants.
Eeach individual olfactory sensory
neuron contains a single type of olfactory receptor that responds
to a limited range of odorant molecules. The axons of cells with
the same receptors converge on a few secondary neurons in the
olfactory bulb, which then can modify the information before sending
it on to the olfactory cortex. The brain uses information from
hundreds of olfactory sensory neurons in different combinations
to create the perception of many different smells, just as combinations
of letters create different words.
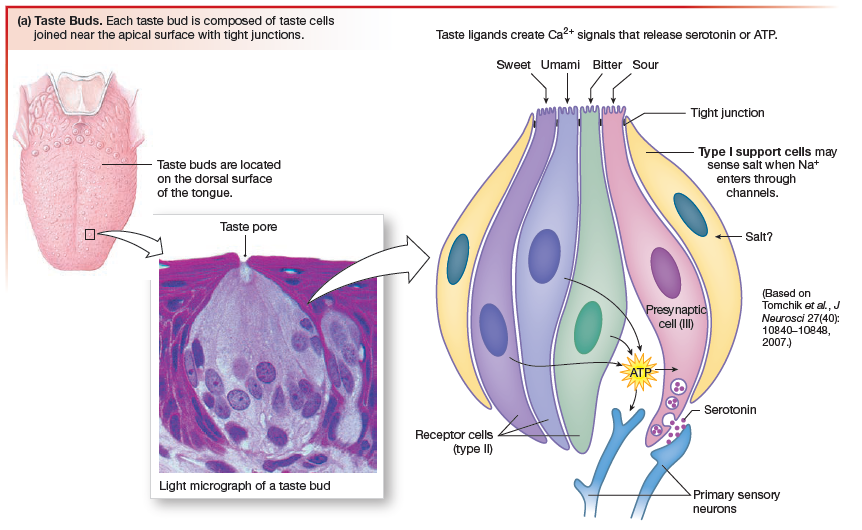
Taste
Taste is currently believed to be a combination of five sensations:
Salty by the presence of Na+ (sodium).
Sweet and umami are associated with nutritious food.
Bitter taste is recognized by the body as a warning of possibly toxic components.
Chemical signals released from taste receptor cells activate primary
sensory neurons (gustatory neurons) whose axons run through cranial nerves VII, IX, and X to the medulla, where they synapse. Sensory information then passes through the thalamus to the gustatory cortex, located in the insula. Central processing of sensory information compares the input from multiple taste receptor cells and interprets the taste sensation based on which populations of neurons are responding most strongly).
Taste buds contain four morphologically distinct cell types designated I, II, and III, plus basal cells (type IV).The type IV basal cells are believed to be precursors of taste receptor cells. Each taste receptor cell is tucked down into the epithelium so that only a tiny tip protrudes into the oral cavity through a taste pore.
- sweet
- sour (acid)
- salty
- bitter
- umami, a taste associated with the amino acid glutamate and some nucleotides.
Salty by the presence of Na+ (sodium).
Sweet and umami are associated with nutritious food.
Bitter taste is recognized by the body as a warning of possibly toxic components.
Chemical signals released from taste receptor cells activate primary
sensory neurons (gustatory neurons) whose axons run through cranial nerves VII, IX, and X to the medulla, where they synapse. Sensory information then passes through the thalamus to the gustatory cortex, located in the insula. Central processing of sensory information compares the input from multiple taste receptor cells and interprets the taste sensation based on which populations of neurons are responding most strongly).
Taste buds contain four morphologically distinct cell types designated I, II, and III, plus basal cells (type IV).The type IV basal cells are believed to be precursors of taste receptor cells. Each taste receptor cell is tucked down into the epithelium so that only a tiny tip protrudes into the oral cavity through a taste pore.
10.4 The Ear: Hearing
The ear is a sense organ that is specialized for two distinct functions:
- hearing
- equilibrium
The ear can be divided into external,
middle, and inner sections, with the neurological elements housed
in and protected by structures in the inner ear.
The vestibular complex of the inner ear is the primary sensor for equilibrium. The
remainder of the ear is used for hearing.
middle, and inner sections, with the neurological elements housed
in and protected by structures in the inner ear.
The vestibular complex of the inner ear is the primary sensor for equilibrium. The
remainder of the ear is used for hearing.
The external ear
The external ear consists of the outer ear, or pinna, and the
ear canal. The pinna funnel sound waves into the ear canal. The ear canal is sealed at its internal end by a thin membranous sheet of tissue called the tympanic membrane, or eardrum.
The middle ear
The tympanic membrane separates the external ear from
the middle ear. The middle ear is an air-filled cavity. Here the Eustachian tube connects with the pharynx to allow middle ear pressure to equilibrate with atmospheric pressure during chewing, swallowing, and yawning.
Three small bones of the middle ear conduct sound from the external environment to the inner ear: the malleus, the incus, and the stapes.
The three bones are connected to one another with the biological equivalent of
hinges. One end of the malleus is attached to the tympanic membrane,
and the stirrup end of the stapes is attached to a thin membrane
that separates the middle ear from the inner ear.
The inner ear
The inner ear consists of two major sensory structures.
- The vestibular apparatus with its semicircular canals is the sensory transducer for our sense of equilibrium.
- The cochlea of the inner ear contains sensory receptors for hearing.
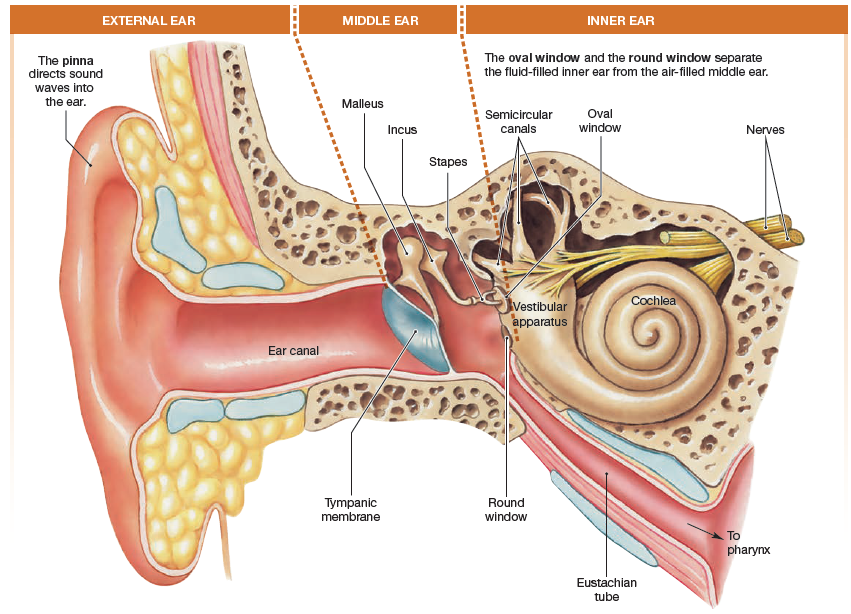
On external view the cochlea is a membranous tube that lies
coiled like a snail shell within a bony cavity. Two membranous
disks, the oval window (to which the stapes is attached) and the
round window, separate the liquid-filled cochlea from the air-filled
middle ear. Branches of cranial nerve VIII, the vestibulocochlear
nerve, lead from the inner ear to the brain.
coiled like a snail shell within a bony cavity. Two membranous
disks, the oval window (to which the stapes is attached) and the
round window, separate the liquid-filled cochlea from the air-filled
middle ear. Branches of cranial nerve VIII, the vestibulocochlear
nerve, lead from the inner ear to the brain.
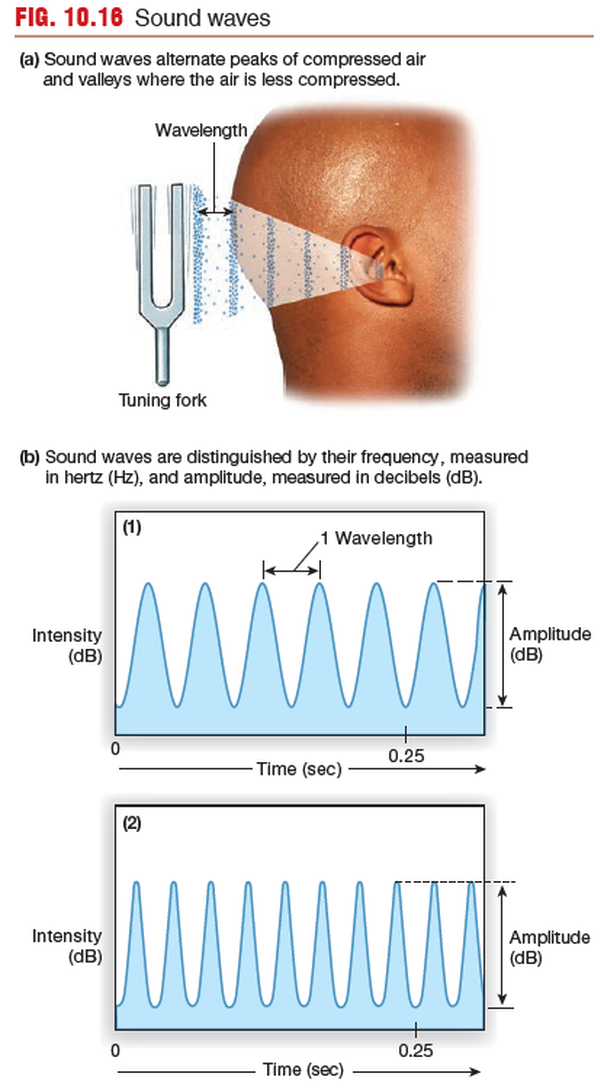
Hearing Is Our Perception of Sound
Hearing is our perception of the energy carried by sound waves,
which are pressure waves with alternating peaks of compressed air
and valleys in which the air molecules are farther apart.
Sound is the brain’s interpretation of the frequency, amplitude,
and duration of sound waves that reach our ears.
Our brains translate frequency of sound waves (the number of wave peaks
that pass a given point each second) into the pitch of a sound.
Low-frequency waves are perceived as low-pitched sounds, such as
the rumble of distant thunder.
The intensity of a sound wave is a function of the wave height, or amplitude.
Intensity is measured on a logarithmic scale in units called decibels
(dB). Each 10-dB increase represents a fold increase in intensity.
which are pressure waves with alternating peaks of compressed air
and valleys in which the air molecules are farther apart.
Sound is the brain’s interpretation of the frequency, amplitude,
and duration of sound waves that reach our ears.
Our brains translate frequency of sound waves (the number of wave peaks
that pass a given point each second) into the pitch of a sound.
Low-frequency waves are perceived as low-pitched sounds, such as
the rumble of distant thunder.
The intensity of a sound wave is a function of the wave height, or amplitude.
Intensity is measured on a logarithmic scale in units called decibels
(dB). Each 10-dB increase represents a fold increase in intensity.
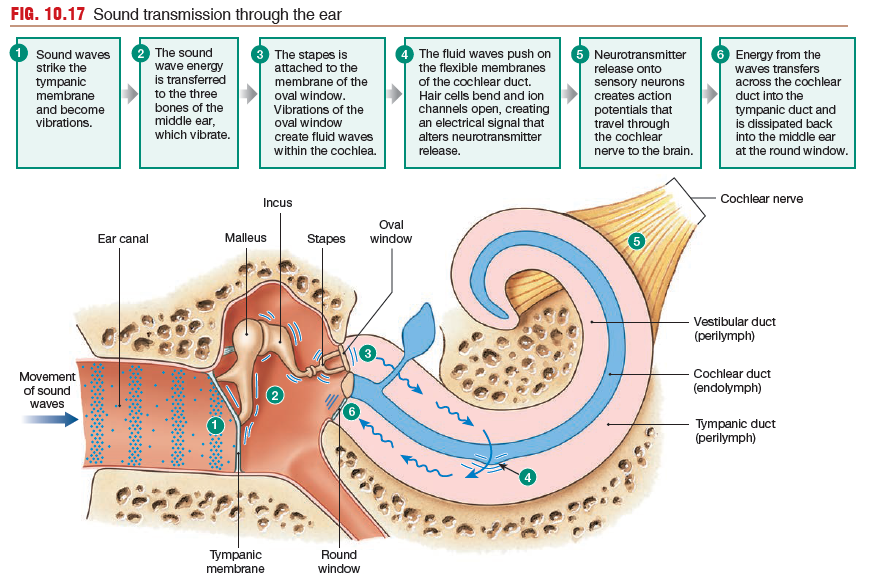
Sound Transduction Is a Multistep Process
Sound waves striking the outer ear are directed down the ear canal until
they hit the tympanic membrane and cause it to vibrate (first transduction).
The tympanic membrane vibrations are transferred to
the malleus, the incus, and the stapes, in that order. The arrangement
of the three connected middle ear bones creates a “lever”
that multiplies the force of the vibration (amplification) so that very
little sound energy is lost due to friction. As the stapes vibrates, it pulls and pushes on the thin tissue of the oval window, to which it is attached. Vibrations at the oval window create waves in the fluid-filled channels of the cochlea (second
transduction).
As waves move through the cochlea, they push on
the flexible membranes of the cochlear duct and bend sensory hair
cells inside the duct. The wave energy dissipates back into the air
of the middle ear at the round window.
Movement of the cochlear duct opens or closes ion channels
on hair cell membranes, creating electrical signals (third transduction).
These electrical signals alter neurotransmitter release (fourth
transduction). Neurotransmitter binding to the primary auditory
neurons initiates action potentials (fifth transduction) that send
coded information about sound through the cochlear branch of the
vestibulocochlear nerve (cranial nerve VIII) and the brain.
they hit the tympanic membrane and cause it to vibrate (first transduction).
The tympanic membrane vibrations are transferred to
the malleus, the incus, and the stapes, in that order. The arrangement
of the three connected middle ear bones creates a “lever”
that multiplies the force of the vibration (amplification) so that very
little sound energy is lost due to friction. As the stapes vibrates, it pulls and pushes on the thin tissue of the oval window, to which it is attached. Vibrations at the oval window create waves in the fluid-filled channels of the cochlea (second
transduction).
As waves move through the cochlea, they push on
the flexible membranes of the cochlear duct and bend sensory hair
cells inside the duct. The wave energy dissipates back into the air
of the middle ear at the round window.
Movement of the cochlear duct opens or closes ion channels
on hair cell membranes, creating electrical signals (third transduction).
These electrical signals alter neurotransmitter release (fourth
transduction). Neurotransmitter binding to the primary auditory
neurons initiates action potentials (fifth transduction) that send
coded information about sound through the cochlear branch of the
vestibulocochlear nerve (cranial nerve VIII) and the brain.
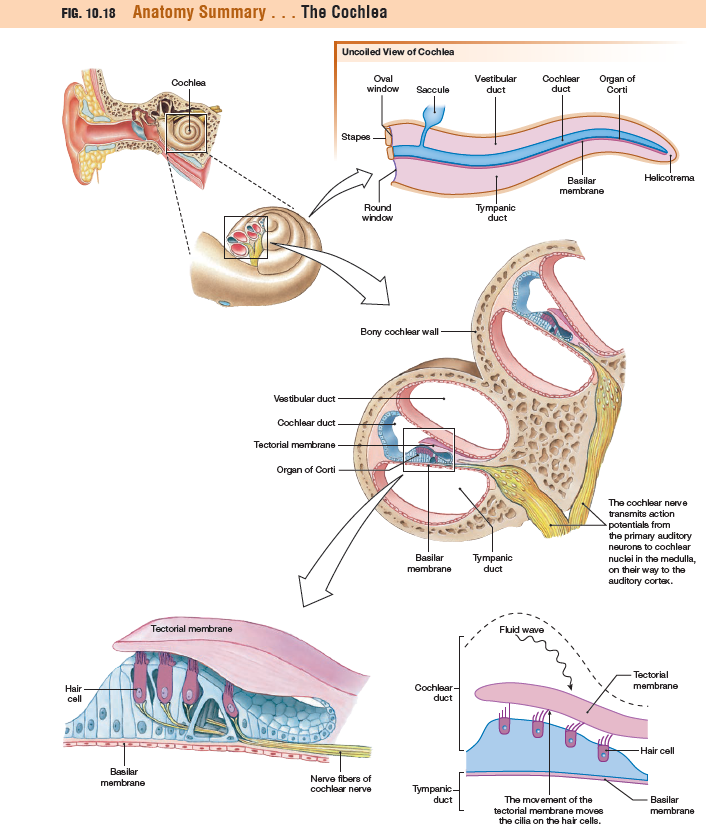
The Cochlea Is Filled with Fluid
The transduction of wave energy into action
potentials takes place in the cochlea of the inner ear.
Uncoiled, the cochlea can be seen to be composed of three parallel, fluid-filled channels:
(1) the vestibular duct, or scala vestibuli
(2) the central cochlear duct, or scala media
(3) the tympanic duct, or scala tympani
The cochlear duct contains the organ of Corti, which is composed
of four rows of hair cell receptors plus support cells. The organ
of Corti sits on the basilar membrane and is partially covered
by the tectorial membrane. These membranes are flexible
tissues that move in response to fluid waves passing through
the vestibular duct. As the waves travel through the
cochlea, they displace basilar and tectorial membranes, creating
up-and-down oscillations that bend the hair cells.
Hair cells, like taste receptor cells, are nonneural receptor
cells. The apical surface of each hair cell is modified into 50–100
stiffened cilia known as stereocilia, arranged in ascending height.
The stereocilia of the hair cells are embedded in the
overlying tectorial membrane. If the tectorial membrane moves,
the underlying cilia do as well.
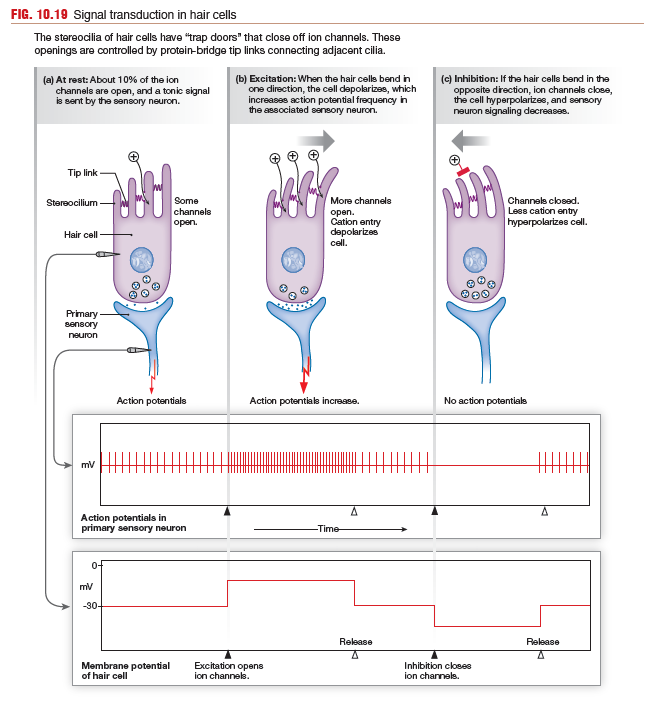
When hair cells move in response to sound waves, their stereocilia
flex, first one way, then the other. The stereocilia are attached
to each other by protein bridges called tip links. The tip links act
like little springs and are connected to gates that open and close
ion channels in the cilia membrane. When the hair cells and cilia
are in a neutral position, about 10% of the ion channels are open,
and there is a low tonic level of neurotransmitter released onto the
primary sensory neuron.
When waves deflect the tectorial membrane so that cilia bend
toward the tallest members of a bundle, the tip links pop more channels
open, so cations (primarily K+ and Ca2+) enter the cell, which then depolarizes. Voltage-gated Ca2+ channels open, neurotransmitter
release increases, and the sensory neuron increases its
firing rate.
When the tectorial membrane pushes the cilia away from
the tallest members, the springy tip links relax and all the ion channels
close. Cation influx slows, the membrane hyperpolarizes, less transmitter
is released, and sensory neuron firing decreases.
The vibration pattern of waves reaching the inner ear is thus
converted into a pattern of action potentials going to the CNS.
Because tectorial membrane vibrations reflect the frequency of the
incoming sound wave, the hair cells and sensory neurons must be
able to respond to sounds of nearly 20,000 waves per second, the
highest frequency audible by a human ear.
potentials takes place in the cochlea of the inner ear.
Uncoiled, the cochlea can be seen to be composed of three parallel, fluid-filled channels:
(1) the vestibular duct, or scala vestibuli
(2) the central cochlear duct, or scala media
(3) the tympanic duct, or scala tympani
The cochlear duct contains the organ of Corti, which is composed
of four rows of hair cell receptors plus support cells. The organ
of Corti sits on the basilar membrane and is partially covered
by the tectorial membrane. These membranes are flexible
tissues that move in response to fluid waves passing through
the vestibular duct. As the waves travel through the
cochlea, they displace basilar and tectorial membranes, creating
up-and-down oscillations that bend the hair cells.
Hair cells, like taste receptor cells, are nonneural receptor
cells. The apical surface of each hair cell is modified into 50–100
stiffened cilia known as stereocilia, arranged in ascending height.
The stereocilia of the hair cells are embedded in the
overlying tectorial membrane. If the tectorial membrane moves,
the underlying cilia do as well.
When hair cells move in response to sound waves, their stereocilia
flex, first one way, then the other. The stereocilia are attached
to each other by protein bridges called tip links. The tip links act
like little springs and are connected to gates that open and close
ion channels in the cilia membrane. When the hair cells and cilia
are in a neutral position, about 10% of the ion channels are open,
and there is a low tonic level of neurotransmitter released onto the
primary sensory neuron.
When waves deflect the tectorial membrane so that cilia bend
toward the tallest members of a bundle, the tip links pop more channels
open, so cations (primarily K+ and Ca2+) enter the cell, which then depolarizes. Voltage-gated Ca2+ channels open, neurotransmitter
release increases, and the sensory neuron increases its
firing rate.
When the tectorial membrane pushes the cilia away from
the tallest members, the springy tip links relax and all the ion channels
close. Cation influx slows, the membrane hyperpolarizes, less transmitter
is released, and sensory neuron firing decreases.
The vibration pattern of waves reaching the inner ear is thus
converted into a pattern of action potentials going to the CNS.
Because tectorial membrane vibrations reflect the frequency of the
incoming sound wave, the hair cells and sensory neurons must be
able to respond to sounds of nearly 20,000 waves per second, the
highest frequency audible by a human ear.
Sounds Are Processed First in the Cochlea
The auditory system processes sound waves so they can be discriminated
by location, pitch, and loudness.
LOCALIZATION - Localization of sound is a complex process that requires sensory input from both ears coupled with sophisticated computation by the brain. The localization of a sound source is an integrative task that requires simultaneous input from both ears. Unless sound is coming from directly in front of a person, it will not reach both ears at the same time. The brain records the time differential for sound arriving at the ears and uses complex computation to create a three-dimensional representation of the sound source.
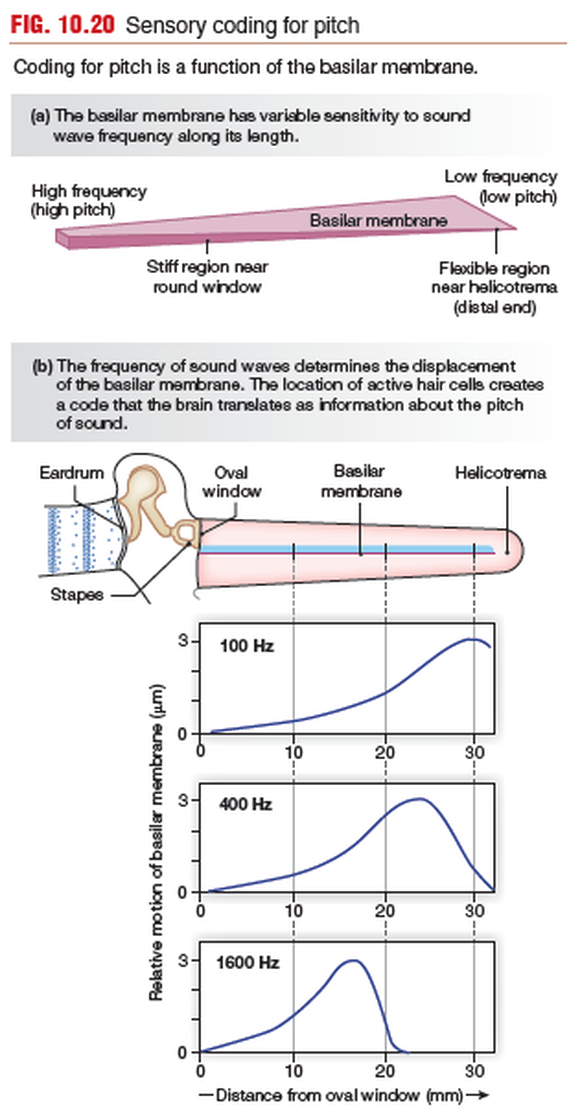
PITCH - Coding sound for pitch is primarily a function of the basilar membrane.
LOUDNESS - Loudness is coded by the ear in the same way that signal strength is coded in somatic receptors. The louder the noise, the more rapidly action potentials fire in the sensory neuron.
by location, pitch, and loudness.
LOCALIZATION - Localization of sound is a complex process that requires sensory input from both ears coupled with sophisticated computation by the brain. The localization of a sound source is an integrative task that requires simultaneous input from both ears. Unless sound is coming from directly in front of a person, it will not reach both ears at the same time. The brain records the time differential for sound arriving at the ears and uses complex computation to create a three-dimensional representation of the sound source.
PITCH - Coding sound for pitch is primarily a function of the basilar membrane.
- High-frequency waves entering the vestibular duct create maximum displacement of the basilar membrane close to the oval window and consequently are not transmitted very far along the cochlea.
- Low-frequency waves travel along the length of the basilar membrane and create their maximum displacement near the flexible distal end.
LOUDNESS - Loudness is coded by the ear in the same way that signal strength is coded in somatic receptors. The louder the noise, the more rapidly action potentials fire in the sensory neuron.
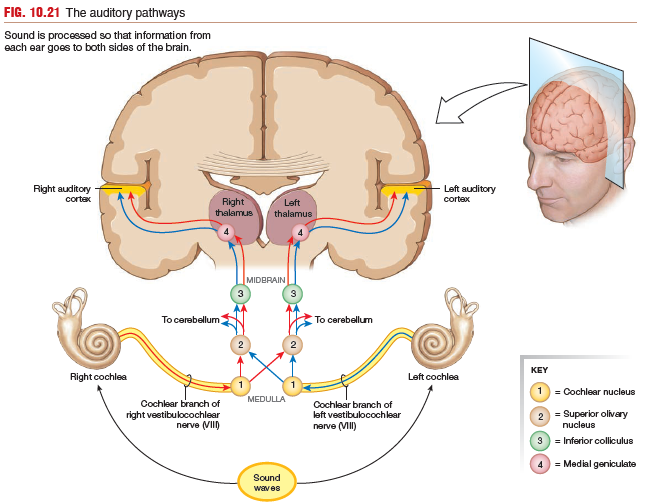
Auditory Pathways Project to the Auditory Cortex
Once the cochlea transforms sound waves into electrical signals,
sensory neurons transfer this information to the brain. The
cochlear (auditory) nerve is a branch of cranial nerve VIII, the vestibulocochlear
nerve.
sensory neurons transfer this information to the brain. The
cochlear (auditory) nerve is a branch of cranial nerve VIII, the vestibulocochlear
nerve.
- Primary auditory neurons project from the cochlea to cochlear nuclei in the medulla oblongata.
- From the medulla, secondary sensory neurons project to two higher nuclei, one ipsilateral (on the same side of the body) and one contralateral (on the opposite side).
- These ascending tracts then synapse in nuclei in the midbrain and thalamus before projecting to the auditory cortex. Collateral pathways take information to the reticular formation and the cerebellum.
10.5 The Ear: Equilibrium
Equilibrium is a state of balance.
The special sense of equilibrium has two components:
another and to the environment.
Visual information also plays an important role in equilibrium, as you know if you have ever gone to one of the 360° movie theaters where the scene tilts suddenly to one side and the audience tilts with it!
Our sense of equilibrium is mediated by hair cells lining the fluid-filled vestibular apparatus of the inner ear. These nonneural receptors respond to changes in rotational, vertical, and horizontal acceleration and positioning. Movement in one direction causes the hair cells to depolarize; with movement in the opposite direction, they hyperpolarize.
The special sense of equilibrium has two components:
- a dynamic component that tells us about our movement through space
- a static component that tells us if our head is not in its normal upright position.
another and to the environment.
Visual information also plays an important role in equilibrium, as you know if you have ever gone to one of the 360° movie theaters where the scene tilts suddenly to one side and the audience tilts with it!
Our sense of equilibrium is mediated by hair cells lining the fluid-filled vestibular apparatus of the inner ear. These nonneural receptors respond to changes in rotational, vertical, and horizontal acceleration and positioning. Movement in one direction causes the hair cells to depolarize; with movement in the opposite direction, they hyperpolarize.
The Vestibular Apparatus Provides Information
about Movement and Position
The vestibular apparatus, also called the membranous labyrinth,
is an intricate series of interconnected fluid-filled chambers.
In humans, the vestibular apparatus consists of two saclike otolith organs—the saccule and the utricle- along with three semicircular canals that connect to the utricle at their bases.
The otolith organs (the saccule and the utricle) tell us about linear acceleration and head position.
The three semicircular canals sense rotational acceleration in various directions.
is an intricate series of interconnected fluid-filled chambers.
In humans, the vestibular apparatus consists of two saclike otolith organs—the saccule and the utricle- along with three semicircular canals that connect to the utricle at their bases.
The otolith organs (the saccule and the utricle) tell us about linear acceleration and head position.
The three semicircular canals sense rotational acceleration in various directions.
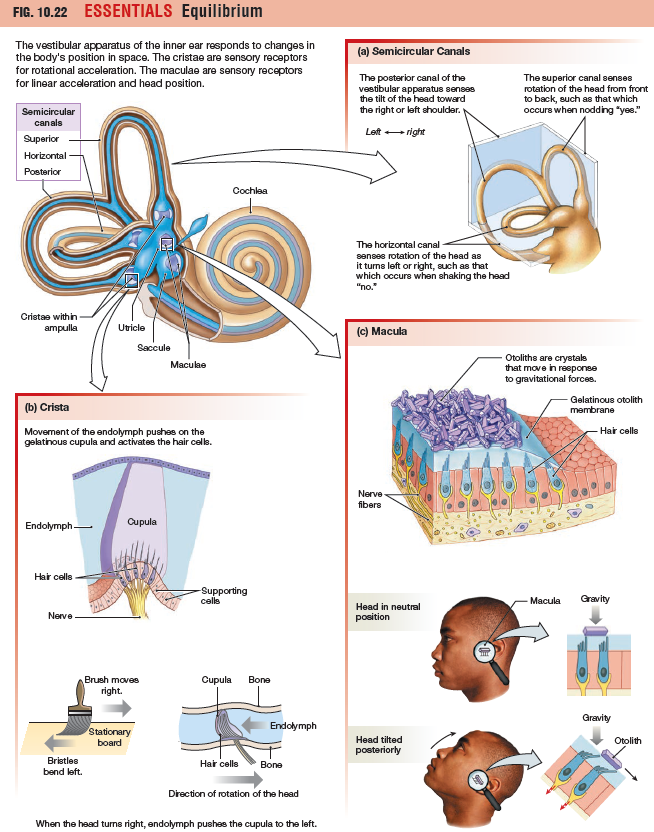
The Semicircular Canals Sense
Rotational Acceleration
The three semicircular canals of the vestibular apparatus monitor
rotational acceleration. They are oriented at right angles to one
another, like three planes that come together to form the corner of
a box. The posterior canal monitors left-to-right rotation, such as the rotation when you tilt your head toward your shoulders or perform a cartwheel. The superior canal is sensitive to forward and back rotation, such as nodding your
head front to back or doing a somersault.
At one end of each canal is an enlarged chamber, the
ampulla, which contains a sensory structure known as a crista.
The crista consists of hair cells and a gelatinous mass, the cupula, that stretches
from floor to ceiling of the ampulla, closing it off. Hair cell cilia are embedded in the cupula.
How is rotation sensed? As the head turns, the bony skull and
the membranous walls of the labyrinth move, but the fluid within
the labyrinth cannot keep up because of inertia (the tendency of a
body at rest to remain at rest). In the ampullae, the drag of endolymph
bends the cupula and its hair cells in the direction opposite
to the direction in which the head is turning.
For an analogy, think of pulling a paintbrush (a cupula attached
to the wall of a semicircular canal) through sticky wet paint (the endolymph)
on a board. If you pull the brush to the right, the drag of the
paint on the bristles bends them to the left (Fig. 10.22b). In the same
way, the inertia of the fluid in the semicircular canal pulls the cupula
and the cilia of the hair cells to the left when the head turns right.
If rotation continues, the moving endolymph finally catches up.
Then if head rotation stops suddenly, the fluid has built up momentum
and cannot stop immediately. The fluid continues to rotate in
the direction of the head rotation, leaving the person with a turning
sensation. If the sensation is strong enough, the person may throw
his or her body in the direction opposite the direction of rotation in a
reflexive attempt to compensate for the apparent loss of equilibrium.
rotational acceleration. They are oriented at right angles to one
another, like three planes that come together to form the corner of
a box. The posterior canal monitors left-to-right rotation, such as the rotation when you tilt your head toward your shoulders or perform a cartwheel. The superior canal is sensitive to forward and back rotation, such as nodding your
head front to back or doing a somersault.
At one end of each canal is an enlarged chamber, the
ampulla, which contains a sensory structure known as a crista.
The crista consists of hair cells and a gelatinous mass, the cupula, that stretches
from floor to ceiling of the ampulla, closing it off. Hair cell cilia are embedded in the cupula.
How is rotation sensed? As the head turns, the bony skull and
the membranous walls of the labyrinth move, but the fluid within
the labyrinth cannot keep up because of inertia (the tendency of a
body at rest to remain at rest). In the ampullae, the drag of endolymph
bends the cupula and its hair cells in the direction opposite
to the direction in which the head is turning.
For an analogy, think of pulling a paintbrush (a cupula attached
to the wall of a semicircular canal) through sticky wet paint (the endolymph)
on a board. If you pull the brush to the right, the drag of the
paint on the bristles bends them to the left (Fig. 10.22b). In the same
way, the inertia of the fluid in the semicircular canal pulls the cupula
and the cilia of the hair cells to the left when the head turns right.
If rotation continues, the moving endolymph finally catches up.
Then if head rotation stops suddenly, the fluid has built up momentum
and cannot stop immediately. The fluid continues to rotate in
the direction of the head rotation, leaving the person with a turning
sensation. If the sensation is strong enough, the person may throw
his or her body in the direction opposite the direction of rotation in a
reflexive attempt to compensate for the apparent loss of equilibrium.
The Otolith Organs Sense Linear Acceleration and Head Position
The two otolith organs, the utricle and saccule, are arranged to sense linear forces. Their sensory structures, called maculae, consist of hair cells, a gelatinous mass known as the otolith membrane, and calcium carbonate and
protein particles called otoliths. The hair cell cilia are embedded in the otolith membrane, and otoliths bind to matrix proteins on the surface of the membrane. If gravity or acceleration cause the otoliths to slide forward or back, the gelatinous otolith membrane slides with them, bending the hair cell cilia and setting off a signal.
The maculae of the utricle sense forward acceleration or deceleration as well as head tilt. In contrast, the maculae of the saccule are oriented vertically when the head is erect, which makes them sensitive to vertical forces, such as dropping downward in an elevator. The brain analyzes the pattern of depolarized and hyperpolarized hair cells to compute head position and direction of movement.
protein particles called otoliths. The hair cell cilia are embedded in the otolith membrane, and otoliths bind to matrix proteins on the surface of the membrane. If gravity or acceleration cause the otoliths to slide forward or back, the gelatinous otolith membrane slides with them, bending the hair cell cilia and setting off a signal.
The maculae of the utricle sense forward acceleration or deceleration as well as head tilt. In contrast, the maculae of the saccule are oriented vertically when the head is erect, which makes them sensitive to vertical forces, such as dropping downward in an elevator. The brain analyzes the pattern of depolarized and hyperpolarized hair cells to compute head position and direction of movement.
10.6 The Eye and Vision
The eye is a sensory organ that functions much like a camera. It
focuses light on a light-sensitive surface (the retina) using a lens
and an aperture or opening (the pupil) whose size can be adjusted
to change the amount of entering light.
Vision is the process through which light reflected from objects in our environment is translated into a mental image. This process can be divided into three steps:
1. Light enters the eye, and the lens focuses the light on the retina.
2. Photoreceptors of the retina transduce light energy into an electrical signal.
3. Neural pathways from retina to brain process electrical signals into visual images.
focuses light on a light-sensitive surface (the retina) using a lens
and an aperture or opening (the pupil) whose size can be adjusted
to change the amount of entering light.
Vision is the process through which light reflected from objects in our environment is translated into a mental image. This process can be divided into three steps:
1. Light enters the eye, and the lens focuses the light on the retina.
2. Photoreceptors of the retina transduce light energy into an electrical signal.
3. Neural pathways from retina to brain process electrical signals into visual images.
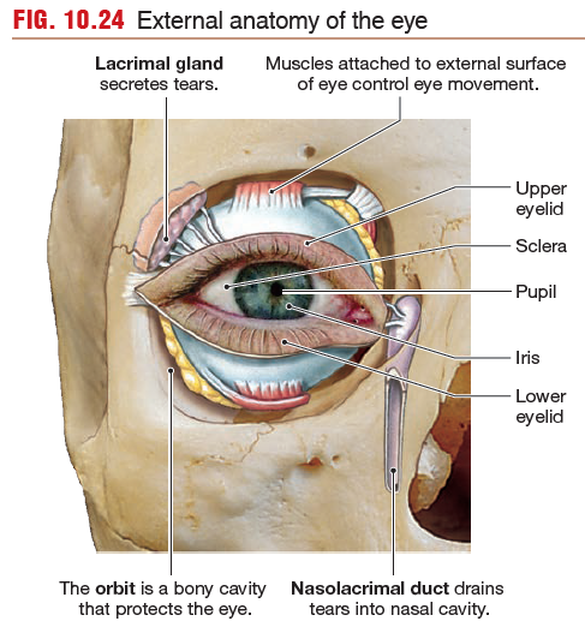
The Skull Protects the Eye
Like sensory elements of the ears, the eyes are protected by a bony cavity,
the orbit, which is formed by facial bones of the skull. Accessory
structures associated with the eye include six extrinsic eye muscles,
skeletal muscles that attach to the outer surface of the eyeball and
control eye movements. Cranial nerves III, IV, and VI innervate
these muscles.
The upper and lower eyelids close over the anterior surface
of the eye, and the lacrimal apparatus, a system of glands
and ducts, keeps a continuous flow of tears washing across the
exposed surface so that it remains moist and free of debris.
The pupil is an opening through which light can pass into
the interior of the eye. Pupil size varies with the contraction and relaxation of a ring of smooth pupillary muscle. The pupil appears
as the black spot inside the colored ring of pigment known as the
iris. The pigments and other components of the iris determine
eye color.
The eye itself is a hollow sphere divided into two compartments
(chambers) separated by a lens. The lens is a transparent disk that focuses light.
The anterior chamber in front of the lens is filled with aqueous
humor.
Behind the lens is a much larger chamber, the vitreous chamber, filled mostly with
the vitreous body that helps maintain the shape of the
eyeball.
The outer wall of the eyeball, the sclera, is composed of
connective tissue.
the orbit, which is formed by facial bones of the skull. Accessory
structures associated with the eye include six extrinsic eye muscles,
skeletal muscles that attach to the outer surface of the eyeball and
control eye movements. Cranial nerves III, IV, and VI innervate
these muscles.
The upper and lower eyelids close over the anterior surface
of the eye, and the lacrimal apparatus, a system of glands
and ducts, keeps a continuous flow of tears washing across the
exposed surface so that it remains moist and free of debris.
The pupil is an opening through which light can pass into
the interior of the eye. Pupil size varies with the contraction and relaxation of a ring of smooth pupillary muscle. The pupil appears
as the black spot inside the colored ring of pigment known as the
iris. The pigments and other components of the iris determine
eye color.
The eye itself is a hollow sphere divided into two compartments
(chambers) separated by a lens. The lens is a transparent disk that focuses light.
The anterior chamber in front of the lens is filled with aqueous
humor.
Behind the lens is a much larger chamber, the vitreous chamber, filled mostly with
the vitreous body that helps maintain the shape of the
eyeball.
The outer wall of the eyeball, the sclera, is composed of
connective tissue.
Light Enters the Eye through the Cornea
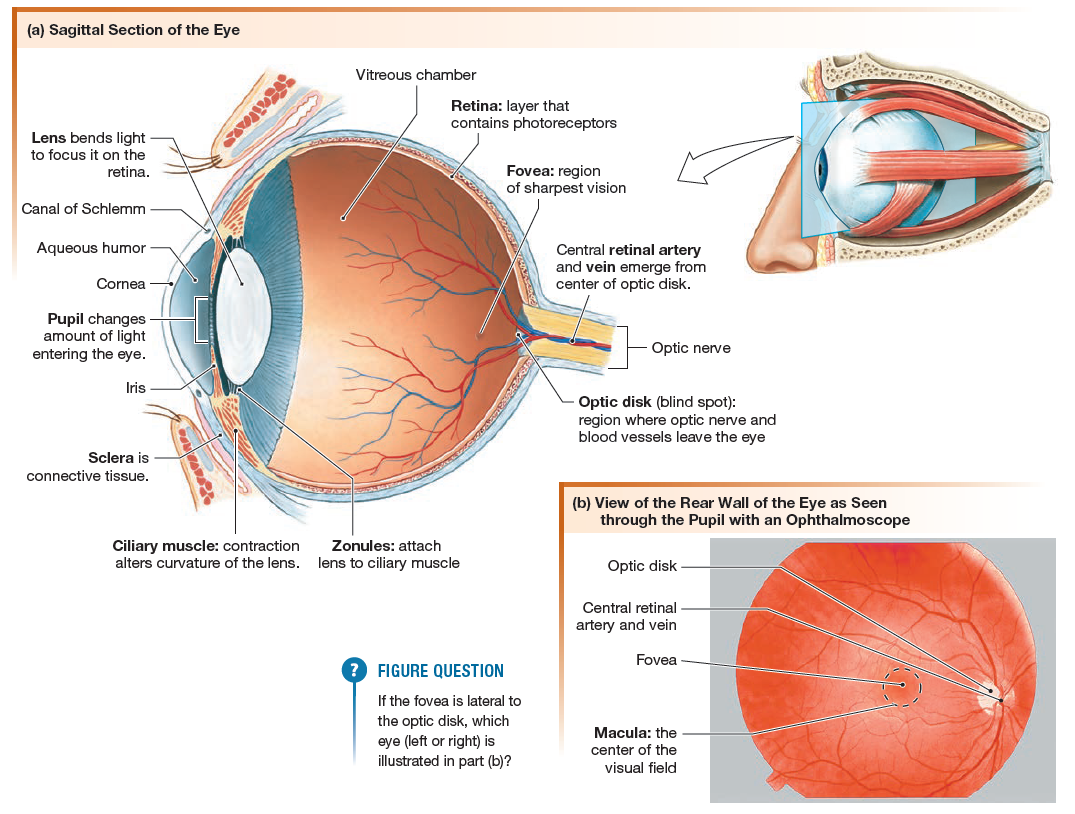
In the first step of the visual pathway, light from the environment
enters the anterior surface of the eye through the cornea,
a transparent disk of tissue that is a continuation of the sclera. This light
is modified two ways.
area.
After passing through the opening of the pupil, light strikes the lens, which has two convex surfaces. The cornea and lens together bend incoming light rays so they focus on the retina, the light-sensitive lining of the eye that contains the photoreceptors.
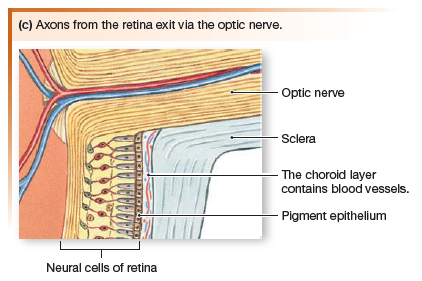
When viewed through the pupil with an ophthalmoscope, the retina is seen to be crisscrossed with small arteries and veins that radiate out from one spot, the optic disk. The optic disk is the location where neurons of the visual pathway form the optic nerve (cranial nerve II) and exit the eye.
Lateral to the optic disk is a small dark spot, the fovea. The fovea and a narrow ring of tissue surrounding it, the macula, are the regions of the retina with the most acute vision.
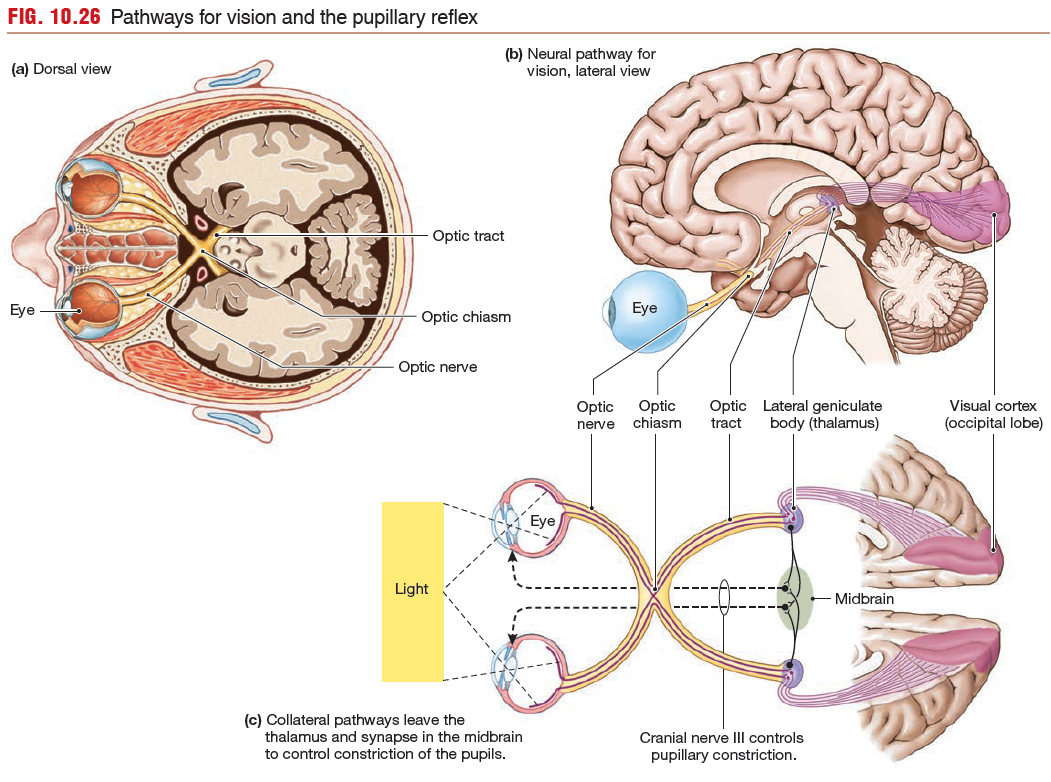
The optic nerves from the retina go to the optic chiasm in the brain, where some of the fibers cross to the opposite side. After synapsing in the lateral geniculate body (lateral geniculate nucleus) of the thalamus, the vision neurons of the tract terminate in the occipital lobe at the visual cortex.
enters the anterior surface of the eye through the cornea,
a transparent disk of tissue that is a continuation of the sclera. This light
is modified two ways.
- First, the amount of light that reaches photoreceptors is modulated by changes in the size of the pupil.
- Second, the light is focused by changes in the shape of the lens.
area.
After passing through the opening of the pupil, light strikes the lens, which has two convex surfaces. The cornea and lens together bend incoming light rays so they focus on the retina, the light-sensitive lining of the eye that contains the photoreceptors.
When viewed through the pupil with an ophthalmoscope, the retina is seen to be crisscrossed with small arteries and veins that radiate out from one spot, the optic disk. The optic disk is the location where neurons of the visual pathway form the optic nerve (cranial nerve II) and exit the eye.
Lateral to the optic disk is a small dark spot, the fovea. The fovea and a narrow ring of tissue surrounding it, the macula, are the regions of the retina with the most acute vision.
The optic nerves from the retina go to the optic chiasm in the brain, where some of the fibers cross to the opposite side. After synapsing in the lateral geniculate body (lateral geniculate nucleus) of the thalamus, the vision neurons of the tract terminate in the occipital lobe at the visual cortex.
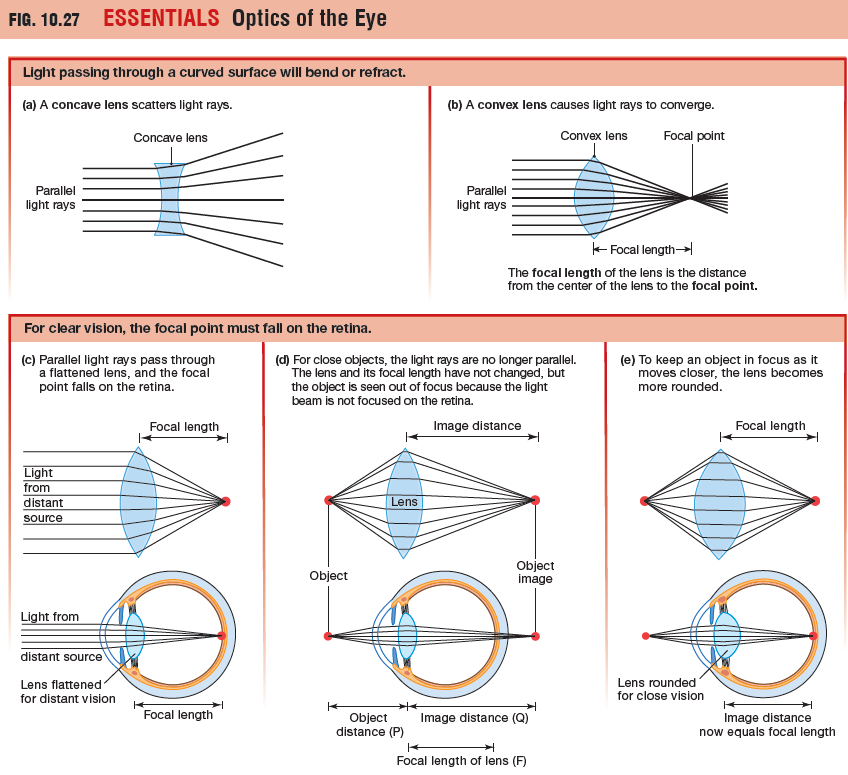
The Lens Focuses Light on the Retina
Light entering the eye is refracted twice:
- when it passes through the cornea
- when it passes through the lens.
The distance from the center of a lens to its focal
point is known as the focal length (or focal distance) of the lens.
When light from an object passes through the lens of the eye,
the focal point and object image must fall precisely on the retina if
the object is to be seen in focus.
Distance Vision - For the normal human eye, any object that is 20 feet or more from the eye creates parallel light rays and will be in focus when the lens is flatter.
Near Vision - When an object is closer than 20 feet to the lens the light rays from the object are not parallel. To keep a near object in focus, the lens must become more rounded to increase the angle of refraction. Making a lens more convex shortens its focal length.
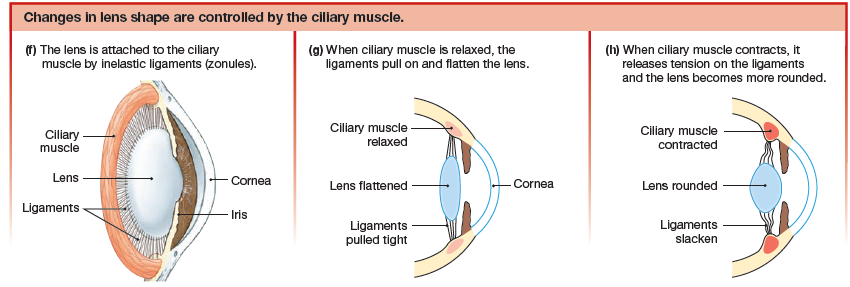
The process by which the eye adjusts the shape of the lens to keep objects in focus is known as accommodation, and the closest distance at which it can focus an object is known as the near point of accommodation. The lens changes shape due to the ciliary muscle which is a ring of smooth muscle that surrounds the lens and is attached to it by the inelastic ligaments called zonules.
- When the ciliary muscle is relaxed, the ring is more open and the lens is pulled into a flatter shape.
- When ciliary muscles contract, the muscle ring gets smaller, releasing tension on the ligaments so that the lens becomes more rounded.
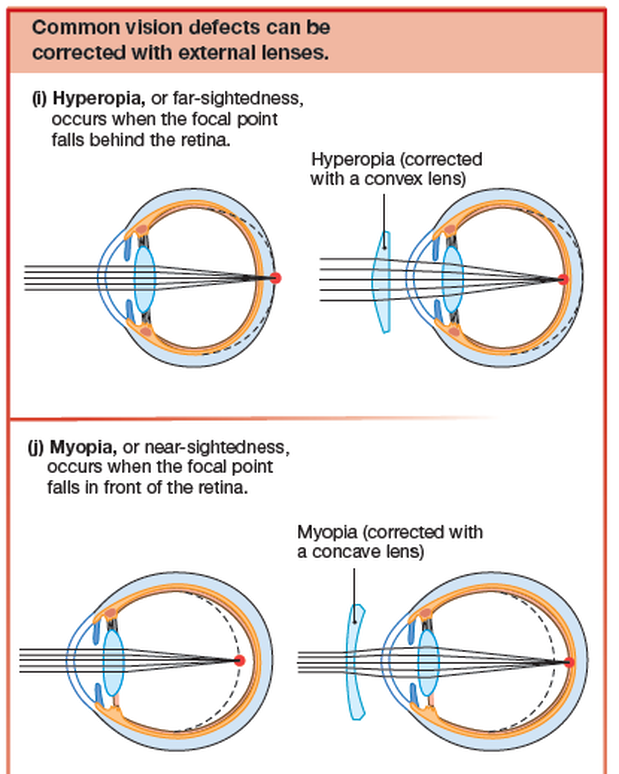
Two other common vision problems are near-sightedness and
far-sightedness.
far-sightedness.
- Near-sightedness, or myopia, occurs when the focal point falls in front of the retina.
- Far-sightedness, or hyperopia, occurs when the focal point falls behind the retina
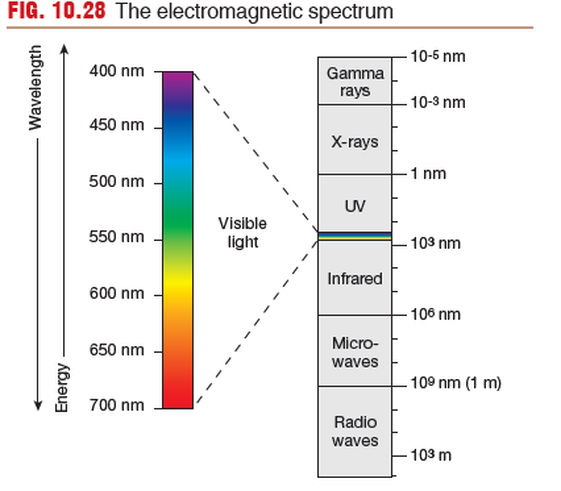
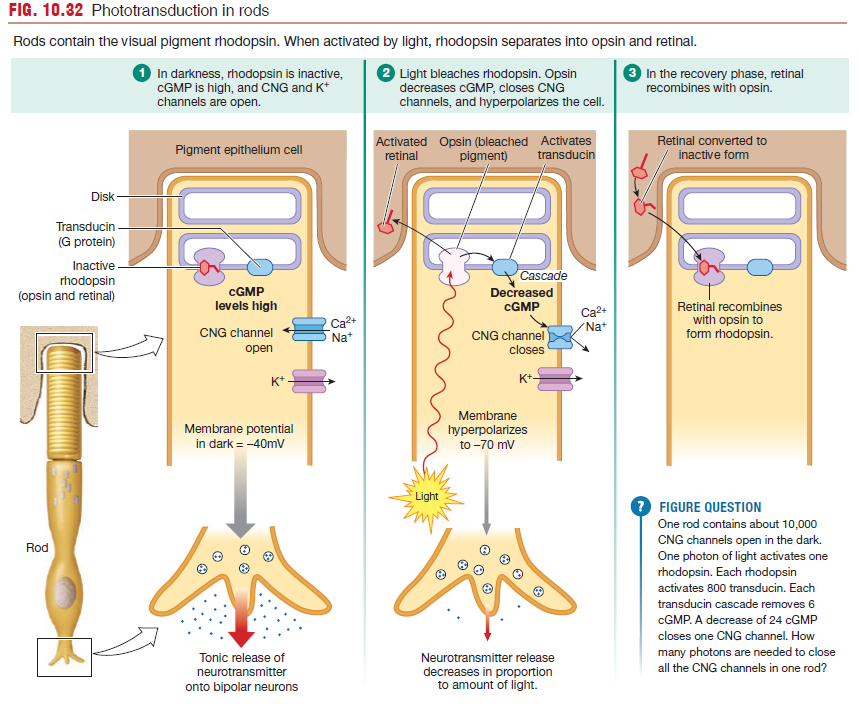
Phototransduction Occurs at the Retina
Once light hits the retina, photoreceptors convert the light energy
into electrical signals. Light energy is part of the electromagnetic
spectrum. For humans, visible light is limited to electromagnetic
energy with waves that have a frequency between 400 and 700nm.
into electrical signals. Light energy is part of the electromagnetic
spectrum. For humans, visible light is limited to electromagnetic
energy with waves that have a frequency between 400 and 700nm.
Phototransduction takes place when light hits the retina, the sensory organ of the eye.
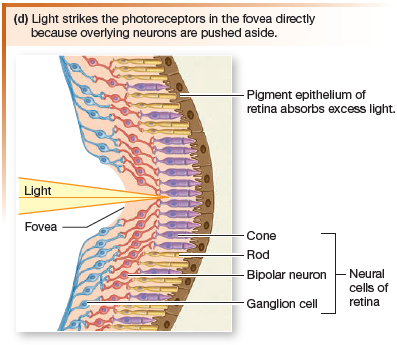
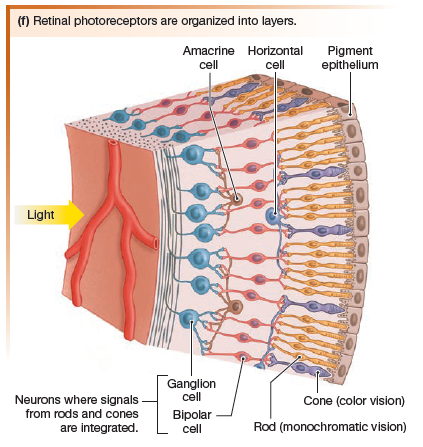
There are five types of neurons in the retinal layers:
The fovea and the macula immediately surrounding it are the areas of most acute vision, and they form the center of the visual field.
When you look at an object, the lens focuses the object image
on the fovea. Light from that section of the visual field falls on the fovea and is in sharp focus.
The image falling on the retina is upside down. Subsequent visual processing by the brain reverses the image again so that we perceive it in the correct orientation.
Sensory information about light passes from the photoreceptors to bipolar neurons, then to a layer of ganglion cells. The axons of ganglion cells form the optic nerve, which leaves the eye at the optic disk. Because the optic disk has no photoreceptors, images projected onto this region cannot be seen, creating
what is called the eye’s blind spot.
There are five types of neurons in the retinal layers:
- photoreceptors
- bipolar cells
- ganglion cells
- amacrine cells
- horizontal cells
The fovea and the macula immediately surrounding it are the areas of most acute vision, and they form the center of the visual field.
When you look at an object, the lens focuses the object image
on the fovea. Light from that section of the visual field falls on the fovea and is in sharp focus.
The image falling on the retina is upside down. Subsequent visual processing by the brain reverses the image again so that we perceive it in the correct orientation.
Sensory information about light passes from the photoreceptors to bipolar neurons, then to a layer of ganglion cells. The axons of ganglion cells form the optic nerve, which leaves the eye at the optic disk. Because the optic disk has no photoreceptors, images projected onto this region cannot be seen, creating
what is called the eye’s blind spot.
Photoreceptors Transduce Light into
Electrical Signals
Photoreceptors are the neurons that convert light energy into electrical signals.
There are two main types of photoreceptors:
There are two main types of photoreceptors:
- rods
- cone
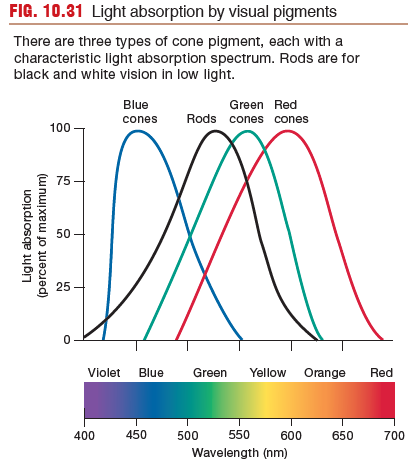
Rods function well in low light and are used in night vision, when objects are seen in black and white rather than in color.
Cones are responsible for high-acuity vision and color vision during
the daytime, when light levels are higher.
The fovea, which is the region of sharpest vision, has a very high density of cones.
Rods and Cones have the same basic structure
(1) an outer segment whose tip touches the pigment epithelium of the retina
(2) an inner segment that contains the cell nucleus and organelles for ATP and protein synthesis
(3) a basal segment with a synaptic terminal that releases glutamate onto bipolar cells.
Cones are responsible for high-acuity vision and color vision during
the daytime, when light levels are higher.
The fovea, which is the region of sharpest vision, has a very high density of cones.
Rods and Cones have the same basic structure
(1) an outer segment whose tip touches the pigment epithelium of the retina
(2) an inner segment that contains the cell nucleus and organelles for ATP and protein synthesis
(3) a basal segment with a synaptic terminal that releases glutamate onto bipolar cells.
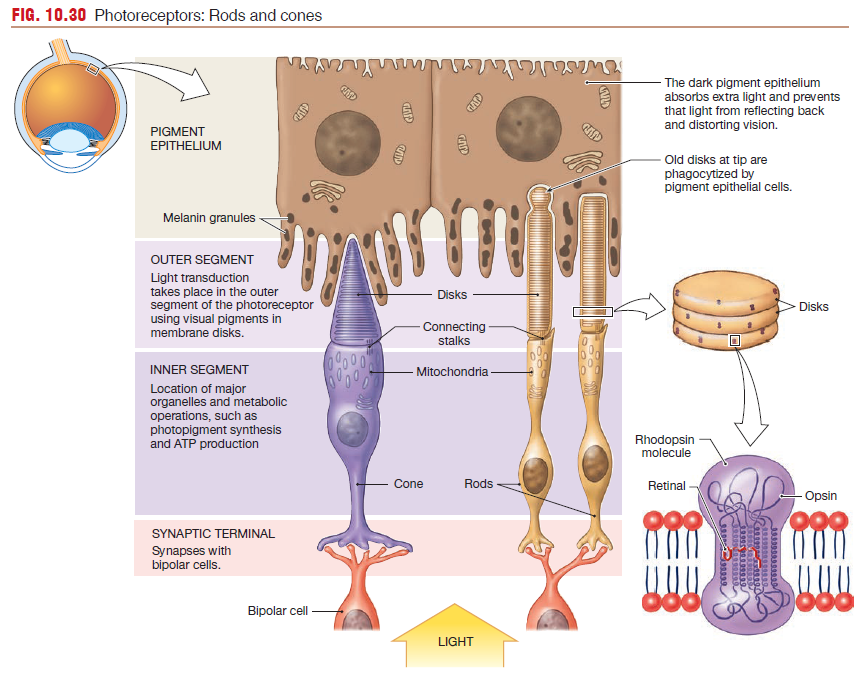
Light-sensitive visual pigments are bound to the disk membranes
in outer segments of photoreceptors.
Red, green, and blue are the three primary colors that make the colors of visible light, just as red, blue, and yellow are the three primary colors that make different colors of paint. The color of any object we are looking at depends on the
wavelengths of light reflected by the object. Green leaves reflect green light, and bananas reflect yellow light. White objects reflect most wavelengths. Black objects absorb most wavelengths, which is one reason they heat up in sunlight while white objects stay cool.
in outer segments of photoreceptors.
- Rods have one type of visual pigment, rhodopsin.
- Cones have three different pigments that are closely related to rhodopsin.
Red, green, and blue are the three primary colors that make the colors of visible light, just as red, blue, and yellow are the three primary colors that make different colors of paint. The color of any object we are looking at depends on the
wavelengths of light reflected by the object. Green leaves reflect green light, and bananas reflect yellow light. White objects reflect most wavelengths. Black objects absorb most wavelengths, which is one reason they heat up in sunlight while white objects stay cool.
There are three types of cone pigment, each with a
characteristic light absorption spectrum. Rods ar e for
black and white vision in low light.
characteristic light absorption spectrum. Rods ar e for
black and white vision in low light.
Signal Processing Begins in the Retina
Visible light causes the photosensitive pigments to alter the membrane potentials of photoreceptors.
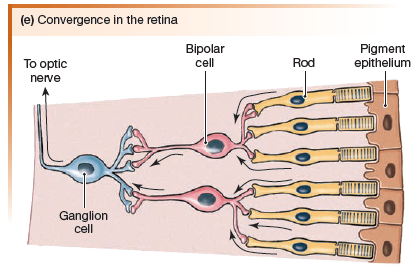
Many photoreceptors may converge onto one single bipolar neuron.
Convergence is minimal in the fovea, where some photoreceptors have a 1:1 relationship with their bipolar neurons. Convergence increases in the peripheral vision at the outer edges of the retina.
Horizontal cells synapse with photoreceptors and bipolar cells
to mediate lateral inhibition in the retina, the same phenomenon
described for touch receptors earlier. Lateral inhibition enhances
stimulus location and contrast.
Amacrine cells modulate information
flowing between bipolar cells and ganglion cells.
Many photoreceptors may converge onto one single bipolar neuron.
Convergence is minimal in the fovea, where some photoreceptors have a 1:1 relationship with their bipolar neurons. Convergence increases in the peripheral vision at the outer edges of the retina.
Horizontal cells synapse with photoreceptors and bipolar cells
to mediate lateral inhibition in the retina, the same phenomenon
described for touch receptors earlier. Lateral inhibition enhances
stimulus location and contrast.
Amacrine cells modulate information
flowing between bipolar cells and ganglion cells.
Bipolar Cells
Glutamate release from photoreceptors onto bipolar
neurons begins signal processing.
There are two types of bipolar
cells:
ON bipolar cells are activated in the light when glutamate secretion by photoreceptors decreases. In the dark, ON bipolar cells are inhibited by glutamate release.
OFF bipolar cells are excited by glutamate release in the dark. In the light, with less glutamate, OFF bipolar cells are inhibited.
By using different glutamate receptors,
one stimulus (light) creates two different responses with a single
neurotransmitter.
Glutamate release from photoreceptors onto bipolar
neurons begins signal processing.
There are two types of bipolar
cells:
- light-on (ON bipolar cells)
- light-off (OFF bipolar cells).
ON bipolar cells are activated in the light when glutamate secretion by photoreceptors decreases. In the dark, ON bipolar cells are inhibited by glutamate release.
OFF bipolar cells are excited by glutamate release in the dark. In the light, with less glutamate, OFF bipolar cells are inhibited.
By using different glutamate receptors,
one stimulus (light) creates two different responses with a single
neurotransmitter.
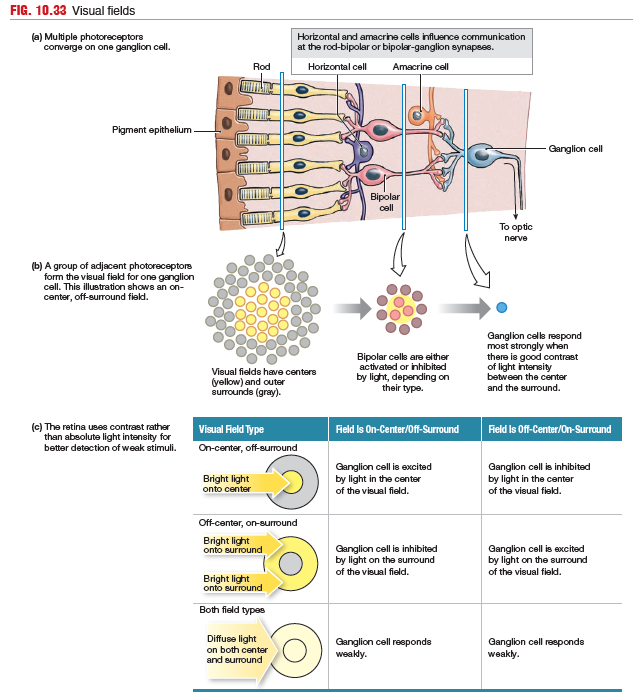
Ganglion Cells
Each ganglion cell receives information from a particular area
of the retina. These areas, known as visual receptive fields. The receptive field of a ganglion cell near the fovea is quite small.
Only a few photoreceptors are associated with each ganglion cell,
and so visual acuity is greatest in these areas. At the edge of the
retina, multiple photoreceptors converging onto a single ganglion
cell results in vision that is not as sharp.
Receptive fields of ganglion cells are roughly circular and are divided
into sections: a round center and its doughnut-shaped surround. This organization allows each ganglion cell to use contrast
between the center and its surround to interpret visual information.
Strong contrast between the center and surround elicits a strong
excitatory response (a series of action potentials) or a strong inhibitory
response (no action potentials) from the ganglion cell. Weak
contrast between center and surround gets an intermediate response.
There are two types of ganglion cell receptive fields.
When light is uniform across a receptive field, the ganglion cell responds weakly. Thus, the retina uses contrast rather than absolute light intensity to recognize objects in the environment. One advantage of using contrast is that it allows better detection of weak stimuli.
of the retina. These areas, known as visual receptive fields. The receptive field of a ganglion cell near the fovea is quite small.
Only a few photoreceptors are associated with each ganglion cell,
and so visual acuity is greatest in these areas. At the edge of the
retina, multiple photoreceptors converging onto a single ganglion
cell results in vision that is not as sharp.
Receptive fields of ganglion cells are roughly circular and are divided
into sections: a round center and its doughnut-shaped surround. This organization allows each ganglion cell to use contrast
between the center and its surround to interpret visual information.
Strong contrast between the center and surround elicits a strong
excitatory response (a series of action potentials) or a strong inhibitory
response (no action potentials) from the ganglion cell. Weak
contrast between center and surround gets an intermediate response.
There are two types of ganglion cell receptive fields.
- on-center/off-surround field - In an on-center/off-surround field, the associated ganglion cell responds most strongly when light is brightest in the center of the field. If light is brightest in the off-surround region of the field, the on-center/off-surround field ganglion cell is inhibited and stops firing action potentials.
- off-center/on-surround field - The reverse happens with off-center/on-surround fields. In an off-center/on-surround field, the associated ganglion cell responds most strongly when light is brightest in the on-surround part the field. If light is brightest in the off-center region of the field, the on-center/off-surround field ganglion cell is inhibited and stops firing action potentials.
When light is uniform across a receptive field, the ganglion cell responds weakly. Thus, the retina uses contrast rather than absolute light intensity to recognize objects in the environment. One advantage of using contrast is that it allows better detection of weak stimuli.
Processing Beyond the Retina
Once action potentials leave ganglion
cell bodies, they travel along the optic nerves to the CNS for
further processing. The optic nerves enter the brain
at the optic chiasm. At this point, some nerve fibers from each eye
cross to the other side of the brain for processing. Some of the information from the right side of each eye’s visual field is processed on the left side of the brain, and information from the left side of the field is processed on the right side of the brain.
cell bodies, they travel along the optic nerves to the CNS for
further processing. The optic nerves enter the brain
at the optic chiasm. At this point, some nerve fibers from each eye
cross to the other side of the brain for processing. Some of the information from the right side of each eye’s visual field is processed on the left side of the brain, and information from the left side of the field is processed on the right side of the brain.
The left visual field of each eye is projected to the visual cortex on
the right side of the brain, and the right visual field is projected to
the left visual cortex. Objects seen by both eyes fall within the
binocular zone and are perceived in three dimensions. Objects
seen with only one eye fall outside the binocular zone and are
perceived in only two dimensions.
the right side of the brain, and the right visual field is projected to
the left visual cortex. Objects seen by both eyes fall within the
binocular zone and are perceived in three dimensions. Objects
seen with only one eye fall outside the binocular zone and are
perceived in only two dimensions.
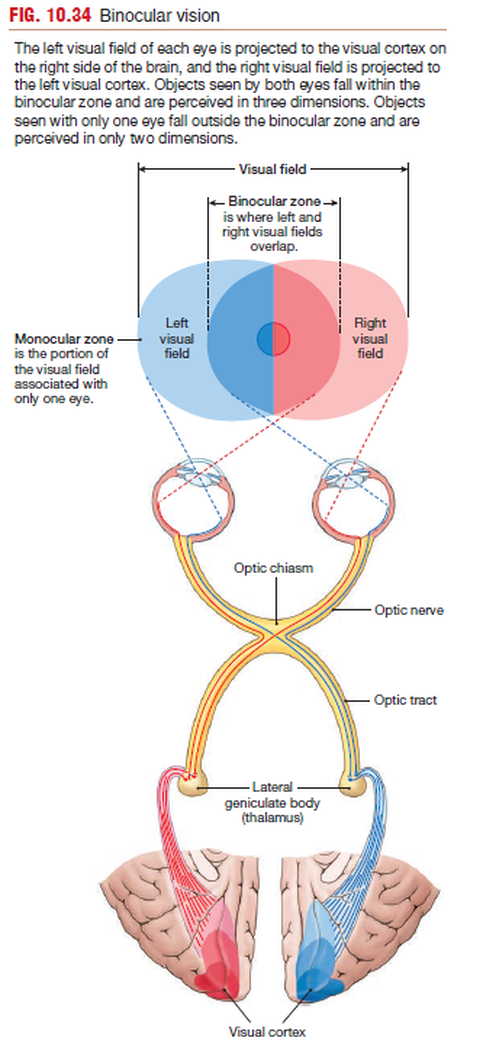
The central portion of the visual field, where left and right
sides of each eye’s visual field overlap, is the binocular zone.
The two eyes have slightly different views of objects in this region,
and the brain processes and integrates the two views to create three-dimensional representations of the objects.
Our sense of
depth perception—that is, whether one object is in front of or
behind another—depends on binocular vision. Objects that fall
within the visual field of only one eye are in the monocular zone
and are viewed in two dimensions.
Once axons leave the optic chiasm, most axons project to the
lateral geniculate body of the thalamus. Here, the optic fibers synapse
onto neurons leading to the visual cortex in the occipital lobe.
The lateral geniculate body is organized in layers that correspond
to the different parts of the visual field, which means that information from adjacent objects is processed together. This topographical
organization is maintained in the visual cortex,
with the six layers of neurons grouped into vertical columns.
Within each portion of the visual field, information is further
sorted by form, color, and movement.
The cortex merges monocular information from the two eyes
to give us a binocular view of our surroundings. Information from
on/off combinations of ganglion cells is translated into sensitivity
to line orientation in the simplest pathways, or into color, movement,
and detailed structure in the most complex. Each of these
attributes of visual stimuli is processed through a separate pathway,
creating a network whose complexity we are just beginning
to unravel.
sides of each eye’s visual field overlap, is the binocular zone.
The two eyes have slightly different views of objects in this region,
and the brain processes and integrates the two views to create three-dimensional representations of the objects.
Our sense of
depth perception—that is, whether one object is in front of or
behind another—depends on binocular vision. Objects that fall
within the visual field of only one eye are in the monocular zone
and are viewed in two dimensions.
Once axons leave the optic chiasm, most axons project to the
lateral geniculate body of the thalamus. Here, the optic fibers synapse
onto neurons leading to the visual cortex in the occipital lobe.
The lateral geniculate body is organized in layers that correspond
to the different parts of the visual field, which means that information from adjacent objects is processed together. This topographical
organization is maintained in the visual cortex,
with the six layers of neurons grouped into vertical columns.
Within each portion of the visual field, information is further
sorted by form, color, and movement.
The cortex merges monocular information from the two eyes
to give us a binocular view of our surroundings. Information from
on/off combinations of ganglion cells is translated into sensitivity
to line orientation in the simplest pathways, or into color, movement,
and detailed structure in the most complex. Each of these
attributes of visual stimuli is processed through a separate pathway,
creating a network whose complexity we are just beginning
to unravel.